4. Genetic diversity in Nordic CWR
4.1 Aims and background
The need for conservation of CWR biodiversity is not limited to the conservation of prioritised species but also comprises conservation of the genetic diversity contained within the species. High levels of within-population genetic diversity facilitate the adaptation to a changing world and is thus important for successful long-term in situ conservation. Conservation of a large amount of genetic diversity is also important from a plant breeding perspective since the usefulness of the preserved diversity is rarely known a priori. Conservation of within-species diversity is thus central for both plant survival and utilisation. Within-species diversity need not be evenly distributed across the range of a species, and rare, unique variants can code for important local adaptations. The distribution of the within-species diversity should ideally also be taken into consideration when planning CWR conservation.
The aim of this part of the project was to gain a better general understanding of the distribution of genetic diversity within selected CWR taxa (within and among populations) in order to more efficiently conserve it for the future, as well as to make species specific recommendation with the same aim.
4.2 CWR species selection and genotyping
As part of the project, we have carried out genetic characterisation of the following six Nordic CWR: Carum carvi L. (caraway); Vaccinium vitis-idaea L. (lingonberry); Schedonorus (Festuca) pratensis (Huds.) P. Beauv. (meadow fescue); Corylus avellana L. (hazel); Fragaria vesca L. (wild strawberry); Vaccinium myrtillus L. (bilberry/European blueberry).
The species were chosen to represent different types of plants (herbs, shrubs, grasses and bushes) and species reproducing both primarily through inbreeding and through outcrossing. Since outreach and education on CWR was an important part of the project, we also prioritised species which we expected to be well-known by the general public and have a clear utility value.
Populations from all species were sampled across their Nordic range and DNA was extracted from multiple individuals per populations. C. carvi, V. vitis-idaea, S. pratensis and C. avellana were genotyped using a Genotyping-by-Sequencing approach while F. vesca and V. myrtillus were genotyped using microsatellite markers.
For the species that were genotyped with the Genotyping by Sequencing approach, C. carvi was sequenced alone while the individuals from the remaining species were combined and sequenced together using a different setup that generates a larger amount of data. Unfortunately, for these species we were unable to retrieve part of the sequencing data, resulting in datasets that contained a high proportion of missing data, i.e. each locus was only genotyped in a subset of the individuals within the species. This makes it difficult to characterise the populations genetically in depth, but we can still make some preliminary conclusions for these species. Genotyping using microsatellites was carried out by visiting internship students. Due to the limited time available for the student who carried out the genotyping, these results are currently incomplete.
4.3 Carum carvi
From C. carvi we genotyped 198 individuals from 16 populations (see Figure 6). Within-population genetic diversity was lowest in a Danish population from Selsø (He = 0.23) and highest in a Norwegian population from Nordmarka, Oslo (He = 0.33). Genetic diversity was neither correlated with latitude nor with longitude and the populations did not show any evidence of inbreeding.
Figure 6. Map showing the locations of the studied Carum carvi populations.
The genetic diversity within the Nordic populations of C. carvi could be divided into three main groups (see Figure 7) and showed an east-westerly distribution. An exception was the northernmost Norwegian population Skjø, which we believe has been a human introduction. The analysis of the Icelandic population Fljótshlíð Hlíðarendi supported the written narrative of C. carvi being originally introduced to Iceland to this location, but with a long history as a naturalised species. Based on the genetic analysis we could make conservation recommendations that would result in conservation of all three genetic clusters and populations with a high level of within-population genetic diversity. We suggested prioritised conservation of the populations Fljótshlíð Hlíðarendi, Iceland; Søndre Ekornholmen, Norway; Äppelö, Åland; Oulanka, Finland and Lebesby Skjøtningberg, Norway.
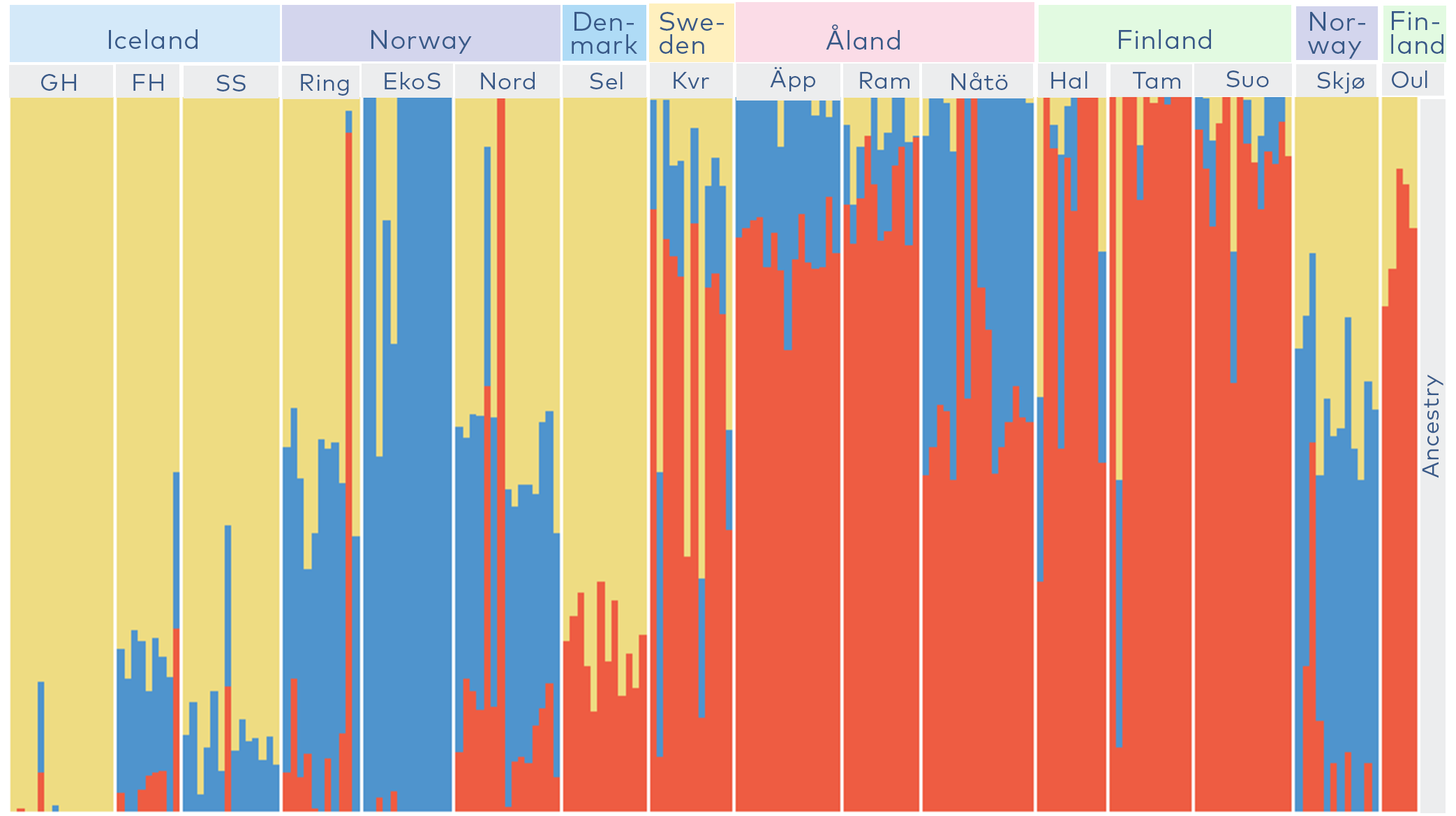
Figure 7. Clustering of the genetic diversity of the studied Carum carvi populations. Each vertical line represents the ancestry of an individual where the different colours show the proportion of the genome of that individual belonging to each of three genetic clusters. White lines separate the studied populations
4.4 Vaccinium vitis-idaea
From V. vitis-idaea we genotyped 284 individuals from 23 populations. The within-population genetic diversity of V. vitis-idaea was somewhat higher than that of C. carvi and with the lowest diversity in the Danish population Melb (He = 0.24) and the highest diversity in Stormossen, Djurvik on Åland (He = 0.40, Figure 8). Icelandic populations tended to contain less within-population diversity than populations from the other Nordic countries. With the exception of a population in northern Sweden, there was little evidence of inbreeding. The data supported a human introduction of some of the Icelandic populations (Rauðavatn, Reykjavik and Þrastarskogur, Suðurland).'
Based on our preliminary analysis, we suggest focusing on the most genetically diverse populations of each country and Åland: Himmelbjerget, Denmark; Oulanka, Finland; Öxnadalur, Iceland; Syltefjord, Norway; Ingarö, Sweden and Stormossen, Åland. This should encompass the full east-western and south-northern ranges of the Nordic region, and the genetic clusters tentatively identified. Additional genotyping of a subset of the populations is currently underway.

Figure 8. Vaccinium vitis-idaea at the collecting site at Stormossen, Djurvik on Åland. Photo: Virva Lyytikäinen.
4.5 Schedonorus (Festuca) pratensis
From S. pratensis we genotyped 115 individuals from eight populations in Finland, Denmark, Norway and Iceland. The within-population genetic diversity was higher for S. pratensis than C. carvi, but similar to that of V. vitis-idaea. The highest diversity was found within a Finnish population from Nuuksio, Lehtimäki (He = 0.38) while the lowest was found in a Danish population from Femmøller (He = 0.32). For several populations, active avoidance of inbreeding seemed to be in place. Analyses of a subset of the data with the genetic loci and individuals with the least amount of missing data, comprising of five populations, tentatively suggested a genetic differentiation between the Icelandic population from Petursey, Suðurland and the other Nordic countries, consistent with an ancient natural colonisation or a human introduction sourced from outside of the Nordic region.
Based on our preliminary analysis, we suggest focusing on the genetically most diverse populations, Nuuksio, Lehtimäki from Finland and Kattrup from Denmark as well as Petursey, Suðurland from Iceland with its differing genetic background. In addition, we recommend further genetic studies to evaluate the genetic diversity of this species in depth.

Figure 9. Schedonorus pratensis at the collecting site at Petursey, Suðurland in Iceland. Photo: Magnus Göransson.
4.6 Corylus avellana
For C. avellana, 178 individuals from 14 populations were genotyped from all the Nordic countries except Iceland, where the species does not grow. Of the four species genotyped with genotyping-by-sequencing, C. avellana had the highest within-population genetic diversity, higher than all three other species. In contrast with the other species, within-population genetic diversity was significantly correlated with latitude and more southern populations contained more genetic diversity than populations from more northern latitudes. The highest within-population diversity was observed in a Danish population from Kattrup (He = 0.42) and the lowest in a Norwegian population from Øyer, Innlandet (He = 0.34). As in S. pratensis, active inbreeding avoidance seemed to occur in some populations.
No clustering of the genetic diversity could be detected with the data available suggesting that conservation of the most genetically diverse populations, regardless of their geographic origin should ensure conservation of a maximum amount of genetic diversity. We tentatively suggest Kattrup and Mols from Denmark, Tønsberg, Færder municipality from Norway, Tvärminne, Hanko from Finland and Ingarö, Värmdö from Sweden (Figure 10) as populations to focus on until more data is available.
No clustering of the genetic diversity could be detected with the data available suggesting that conservation of the most genetically diverse populations, regardless of their geographic origin should ensure conservation of a maximum amount of genetic diversity. We tentatively suggest Kattrup and Mols from Denmark, Tønsberg, Færder municipality from Norway, Tvärminne, Hanko from Finland and Ingarö, Värmdö from Sweden (Figure 10) as populations to focus on until more data is available.

Figure 10. Corylus avellana at the collection site at Ingarö, Värmdö in Sweden. Photo: Anna Palmé.
4.7 Fragaria vesca
For F. vesca 95 individuals from nine populations were genotyped (for an example see Figure 11). As in C. avellana, genetic diversity decreased with latitude with the highest level of diversity in a Danish population from Kattrup and the lowest in an Icelandic population from Ásbyrgi. Our data tentatively suggested the presence of three genetic groups and conservation efforts should hence ideally include the most diverse populations from each group. Additional genotyping is currently underway and until the results from these become available the Icelandic population from Borgarfjörður, the Danish population from Kattrup and the Finnish population from Nåtö on Åland are the most interesting populations from a conservation perspective.

Figure 11. Fragaria vesca growing in the collection site in Abisko in Sweden. Photo: Mora Aronsson.
4.8 Vaccinium myrtillus
For V. myrtillus, 93 individuals from six populations from Finland, Iceland and Sweden were genotyped. The results suggested the highest levels of within-population diversity in a Finnish population from Nuuksio and the lowest in a Swedish population from Abisko (Figure 12). High levels of inbreeding were suggested for the Finnish population from Oulanka. With the available data the different populations cannot be grouped into any genetic clusters, suggesting the identity of a population may be of lesser importance than its genetic diversity. Additional genotyping is currently underway.

Figure 12. Collection of Vaccinium myrtillus in Abisko, Sweden. Photo: Mora Aronsson.