The Barents Sea
Geir Ottersen, Institute of Marine Research, Norway and
Thorsten Blenckner, Stockholm Resilience Centre, Sweden
Thorsten Blenckner, Stockholm Resilience Centre, Sweden
The effects of climate change are of particular importance in the Arctic due to the temperature increase there, resulting in a fundamental change from a seasonally ice-covered to a permanently open ocean system. The northern and eastern parts of the Barents Sea (BS) are essentially of arctic nature. They have been, and to a large degree still are, seasonally ice-covered, with a long and dark Arctic winter and strongly light limited production. The southwestern part of the BS is boreal, but the boundary between the two regimes is not locked into latitude or longitude but shifts with the water masses (Fig. 11).

Figure 11. An artist’s view of the southern Barents Sea. Credit: Institute of Marine Research, Norway.
As briefly described earlier, by absorbing large quantities of CO2 from the atmosphere, ocean chemistry is changing and especially the surface layer is becoming more acidic (IPCC, 2019). Ocean acidification may become a threat by affecting fish eggs and larvae or through phytoplankton, but too little is yet known to draw any conclusions. Lack of oxygen is generally not considered to be a threat in the open Barents Sea.
Primary Production
Primary production, mainly phytoplankton, lays the foundation for much of marine life. How climate change affects this production will be important for everything from zooplankton to fish and whales. Primary production depends on nutrients reaching the upper layers where there is light. Thus, future production depends a lot on the availability of nutrients, which is regulated by the degree of the stratification of the sea and wind conditions. Strong stratification hinders nutrients entering the upper photic zone, thus reducing primary production. On the other hand, increased windiness will reduce the layering, mix up water from below and increase the nutrient levels in the upper layer. At high latitudes a large proportion of the yearly primary production is coupled to the phytoplankton bloom happening when the sea-ice retreats in the spring. These areas now undergo major changes related to diminishing ice coverage, changes that are expected to strengthen with further climate change. A future (more or less) ice-free BS will likely lead to significant changes in spring bloom dynamics and seasonality more generally, total productivity, and ecosystem structure (Sandø et al. 2021). Retreating ice will in large areas drive a transition from a system dominated by ice-algal production to one dominated by open-water phytoplankton blooms with an overall marked decrease in ice algae and increase in open-water phytoplankton production (Mueter et al. 2021). However, the development is complicated as the expected warming of surface seawater and increased freshwater from precipitation and (for a period) ice-melt leads to stronger stratification, while, acting to the contrary, stronger winds and increased storminess can have a positive effect on plankton production through weaker layering and thus increased upward mixing of nutrients.
Further, significant positive relationships have been found between ice-free conditions (open water area and duration) and net primary production (NPP; Arrigo and Donald 2020). The estimated annual NPP for the Barents Sea more than doubled over the 1998–2017 period, from around 40 to over 100 Tg C (teragrams, million tonnes of carbon); Dalpadado et al., 2020). The strong increase in NPP is the result of reduction of sea ice, extending both the area and period available for phytoplankton production. Over this ca 20-year period, the spring and summer phytoplankton blooms expanded and shifted to the north and east (Dalpadado et al. 2020).
In areas where ice extent has decreased, satellite-derived measurements of chlorophyll a show that the timing of the peak spring phytoplankton bloom has advanced by over a month. Using satellite ocean colour data, Renaut et al. (2018) observed a significant increase in primary productivity of phytoplankton spring blooms in the Barents Sea. A northward expansion of these blooms at a rate of 1° per decade driven by the Barents and the Kara regions was also detected. Satellite-based results confirm information from other sources that phytoplankton dynamics in the Barents Sea ecosystem have been changing rapidly and that this is driven mainly by bottom-up climatic processes (Renaut et al. (2018, Dalpadado et al. 2020).
The study by Sandø et al. (2021) examined how future variability and trends in temperature, sea-ice extent, light and wind-induced mixing in the BS will affect the lower trophic levels in the marine ecosystem. They combined downscaling of the NorESM1-M earth system model for the IPCC RCP4.5 scenario by means of the Regional Ocean Model System (ROMS) with the coupled physical–chemical–biological model system NORWegian ECOlogical Model system end-to-end, NORWECOM.E2E (Aksnes et al. 1995, Skogen and Søiland 1998). They projected that during the period 2010-2070 the northernmost parts of the BS gained increased access to light during the productive season due to decreased sea-ice extent, leading to increased primary and secondary production in periods of low sea ice concentrations. In the southern parts, variable access to nutrients as a function of wind-induced mixing and mixed layer depth were found to be the most dominating factors controlling variability in primary production (Sandø et al. 2021).
Projections by NORWECOM.E2E and evaluations by an expert panel were further used to project future primary production (and other variables) in the Barents Sea by Sandø et al. (2024). They estimated accumulated directional effects of climate change on primary production during 2015-2100 as a function of climate exposure and sensitivity attributes under the three IPCC climate scenarios SSP1-2.6 (low emission), SSP2-4.5 (moderate emission), and SSP5-8.5 (extremely high emission; Fig. 12).
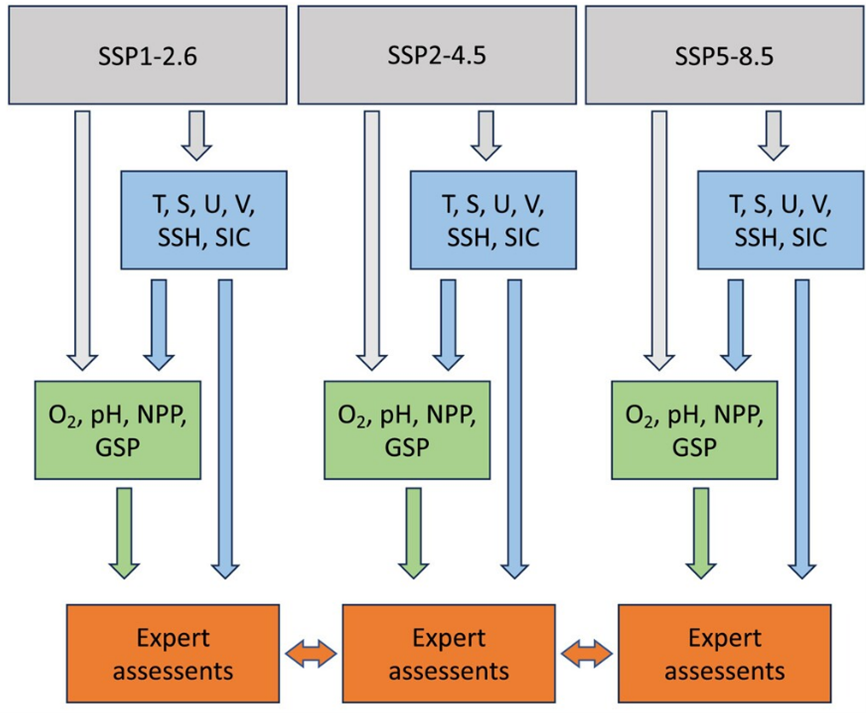
Figure 12. Rough outline of the approach behind the assessments of climate change impact on plankton and fish undertaken by Sandø et al. (2024). The Norwegian Earth System Model (NorESM) produces atmospheric forcing and boundary conditions for ocean downscaling within each of three scenarios (upper grey boxes). Regionally downscaled scenarios from the regional ocean model NEMO-NAA10km produces hydrography and ocean currents, sea surface height and sea ice extent within each of the same scenarios (blue boxes). The ecosystem model NORWECOM.E2E produces biogeochemical variables for each scenario (green boxes). Finally, based on the above and knowledge on selected sensitivity attributes expert assessments are conducted to estimate accumulated directional effects on plankton and a wide selection of fish (bottom orange boxes). From Sandø et al. (2024).
Sandø et al. (2024) evaluated four parts of the BS separately, Polar (coldest and most northerly), Arctic, Atlantic, and Coastal. For SSP2-4.5 and even more so in SSP5-8.5 an increase in net primary production was found for the Polar, Arctic, and Coastal regions. Within SSP1-2.6 the projected changes were very small for all four regions, as were they for the Atlantic region within all scenarios (Sandø et al. 2024). Results are here shown for the Arctic and Atlantic parts of the BS (Fig. 13).
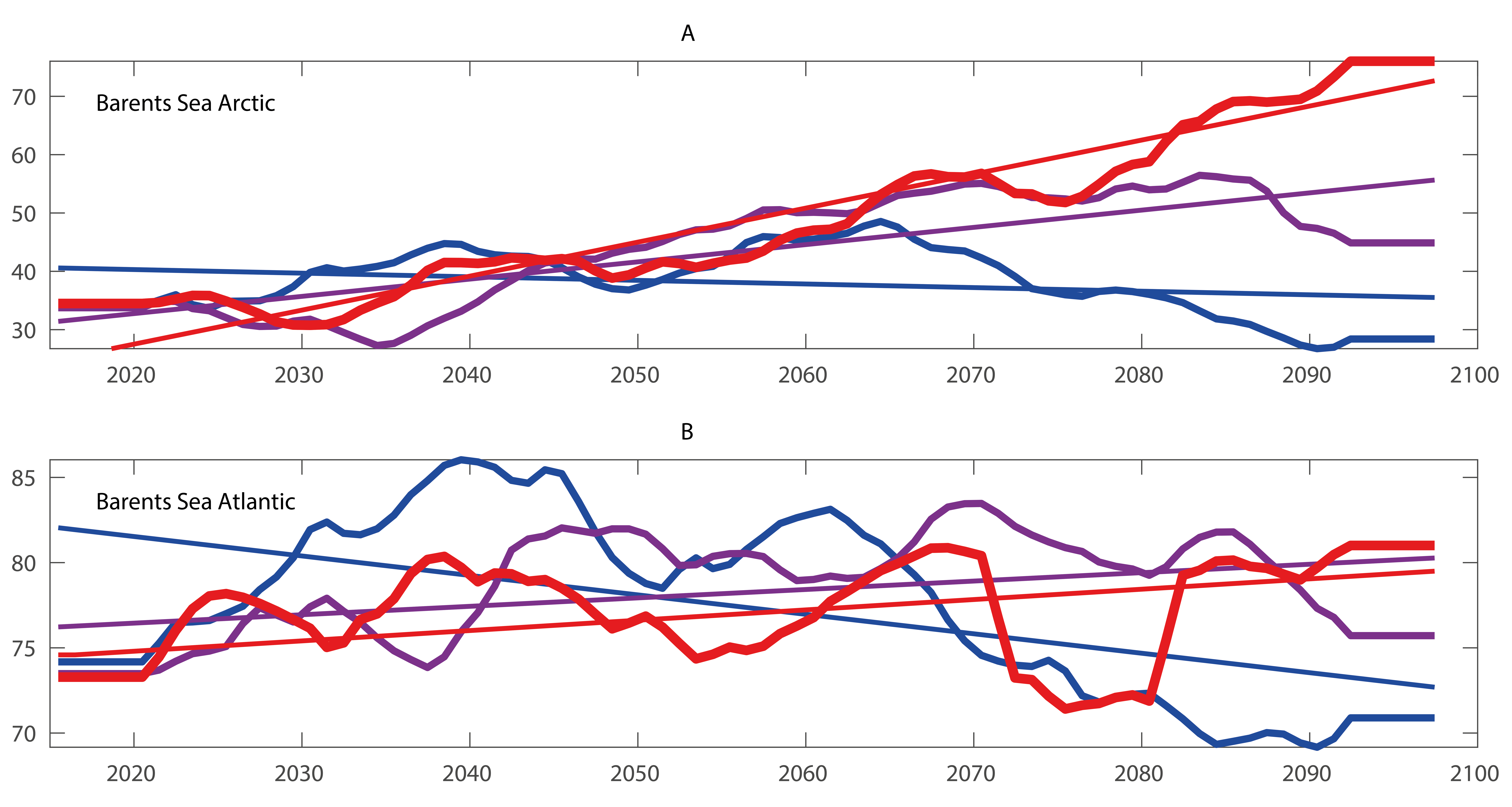
Figure 13. Projected annually averaged Net Primary Production in Tg C (y-axes) in the Barents Sea for the period 2015–2100 in (A) Arctic, and (B) Atlantic region. Coloured time series indicate the scenarios SSP1-2.6 (blue), SSP2-4.5 (purple), and SSP5-8.5 (red), and corresponding straight lines their trends.
Secondary production - zooplankton
In the Northern, seasonally ice-covered, part of the Barents Sea, primary production has been strongly light limited. A short and geographically limited phytoplankton bloom has also limited the zooplankton potential to utilize the existing particulate organic material due to low (phytoplankton) prey density. Future decrease in ice coverage will increase available light and expand and prolong the secondary production significantly as compared to the present. This change is particularly pronounced under the SSP8.5 scenario. For the polar region of the BS the annually averaged April–July Gross secondary production (GSP) has been projected to increase by on average more than 50 g/cm2 over the period 2015–2100, in some smaller areas even more. Also for the Arctic and Atlantic regions the projected changes under the SSP8.5 scenario are pronounced (Fig. 14; Sandø et al. 2024).

Figure 14. Projected annually averaged Gross Secondary Production (in g/cm2; y-axes) in the Barents Sea for the period 2015-2100 in (A) Arctic, and (B) Atlantic region. Coloured time series indicate the scenarios SSP1-2.6 (blue), SSP2-4.5 (purple), and SSP5-8.5 (red), and corresponding straight lines their trends.
The transition from ice-algae to open water plankton blooms also affects the size and species composition of zooplankton communities, with a shift from large-bodied and typically lipid-rich zooplankton, whose early life stages are associated with the ice, to smaller and often less nutritious zooplankton species in the absence of ice (Aarflot et al. 2018, Mueter et al. 2021). Such changes in the size structure of lower trophic levels have been supported by laboratory studies and models.
In the western part of the Barents Sea, Aarflot et al (2018) observed indications of borealization of the zooplankton community, with a decreasing proportion of the Arctic C. glacialis from around year 2000 onwards and increase of the smaller boreal/North Atlantic C. finmarchicus during the same period. This is likely linked to increasing temperatures in the area and the pattern may strengthen with future more permanent warming.
Such species replacement within the Calanus complex has caused concern for ecosystem functioning and energy transfer to higher trophic levels. Calanus species store large quantities of lipids, making these zooplankton a critical link in marine food-webs. The Arctic Calanus species are usually larger and, importantly, have been suggested to contain disproportionately larger lipid stores than their boreal congeners. As pointed to above, there are already shifts from the larger, lipid-rich Arctic species C. glacialis and C. hyperboreus, toward C. finmarchicus in the European Arctic, including the Barents Sea. Continued climate warming and subsequent changes in primary production regimes are expected to strengthen this development. It has been argued by several authors that there will be severe negative consequences, e.g., Mueter et al (2021) state that this will have detrimental effects on fishes that depend on high-lipid prey for overwinter survival as well as seabirds. However, this is debated. Renaud et al. (2018) suggest that lipid content is closely related to body size for all three species. Following this line of argument, lipid content is not a species-specific trait, and further, there is considerable overlap in size between C. finmarchicus and C. glacialis. This suggests that climate driven shifts in dominant Calanus species may not negatively impact their consumers in the Barents Sea.
Benthic plants and animals
The bottom-dwelling organisms in the Barents Sea have adapted to the temperatures in their living areas. Species in the southwest are thus adapted to warmer Atlantic climate and will be able to spread northwards with expanding Atlantic water masses. Contrary, Arctic species in the northeast may become more vulnerable, especially species that prefer stable cold temperatures. This is similar as for Barents Sea fish, but benthic animals are typically sessile species and individuals cannot swim away from warmer water. The populations can move, but only gradually through, for example, larval dispersal.
Further, in connection with the expected increasing sea temperatures sea ice will retreat northwards. This will lead to expansion or movement of the distribution not only of commercially targeted fish species, but also fisheries. Therefore, bottom living animals and plants in areas that were previously unaffected, may now come under pressure from fisheries, especially bottom trawling (Miljøstatus 2024).
Some bottom-oriented animals can indeed move actively, like crabs. The red king crab was introduced to the Barents Sea during the 1960 as a consequence of a Russian initiative to establish a new fishable resource. The crabs have since their release locally in the Murmansk fjord expanded to cover most of the near-coastal zone of the Barents Sea. No clear link between this development and changes in climate have so far been detected. However, it has been suggested that with increased warming the king crab may spread northwards and further offshore from its current near-coastal distribution (Christiansen et al. 2015).
The other numerous and expanding crab species in the Barents Sea is the snow crab. Although some observations were made from the mid-1990s onwards, snow crab is reckoned to be established as a species in the Barents Sea from around 2005. Their numbers have since increased significantly (Lorentzen et al. 2018). This is not clearly linked to climate. It has earlier been argued that future spreading of the snow crab in the Barents Sea will depend on warming (Pavlov and Sokolov 2003, Bakanev 2015). Mueter et al (2021) investigated a broad range of future scenarios for Subarctic and Arctic marine ecosystems. They envisage that snow crab will contract in the Subarctic but increase on Arctic shelves, the Barents Sea included in the latter. They argue that higher temperatures, combined with increased predation from expanding predator populations, reduces the survival of early stages of snow crab at the southern end of their current distribution. However, longer ice-free season and warming in more northern regions create favourable conditions for growth (Mueter et al. 2021). This is in accordance with Tao et al. (2024), who examined the impact of climate change on snow crabs and their fishery, including in the Barents Sea. They point to Arctic sea ice extent as a crucial climate factor affecting snow crab biomass and harvests.
Although northern prawns spend significant time in the water column, they are generally considered benthic. In their projections for the Barents and Norwegian Sea with the NoBa Atlantis ecosystem model, Nilsen et al. (2024) examined the directional response of a range of species to the three climate scenarios SSP1-2.6 (low), SSP2-4.5 (intermediate), and SSP5-8.5 (extremely high). Of all the species the prawns were found to have the most positive response to the strong warming under SSP5-8.5 but also responded more positively to SSP2-4.5 as compared to SSP1-2.6 (Fig. 15).
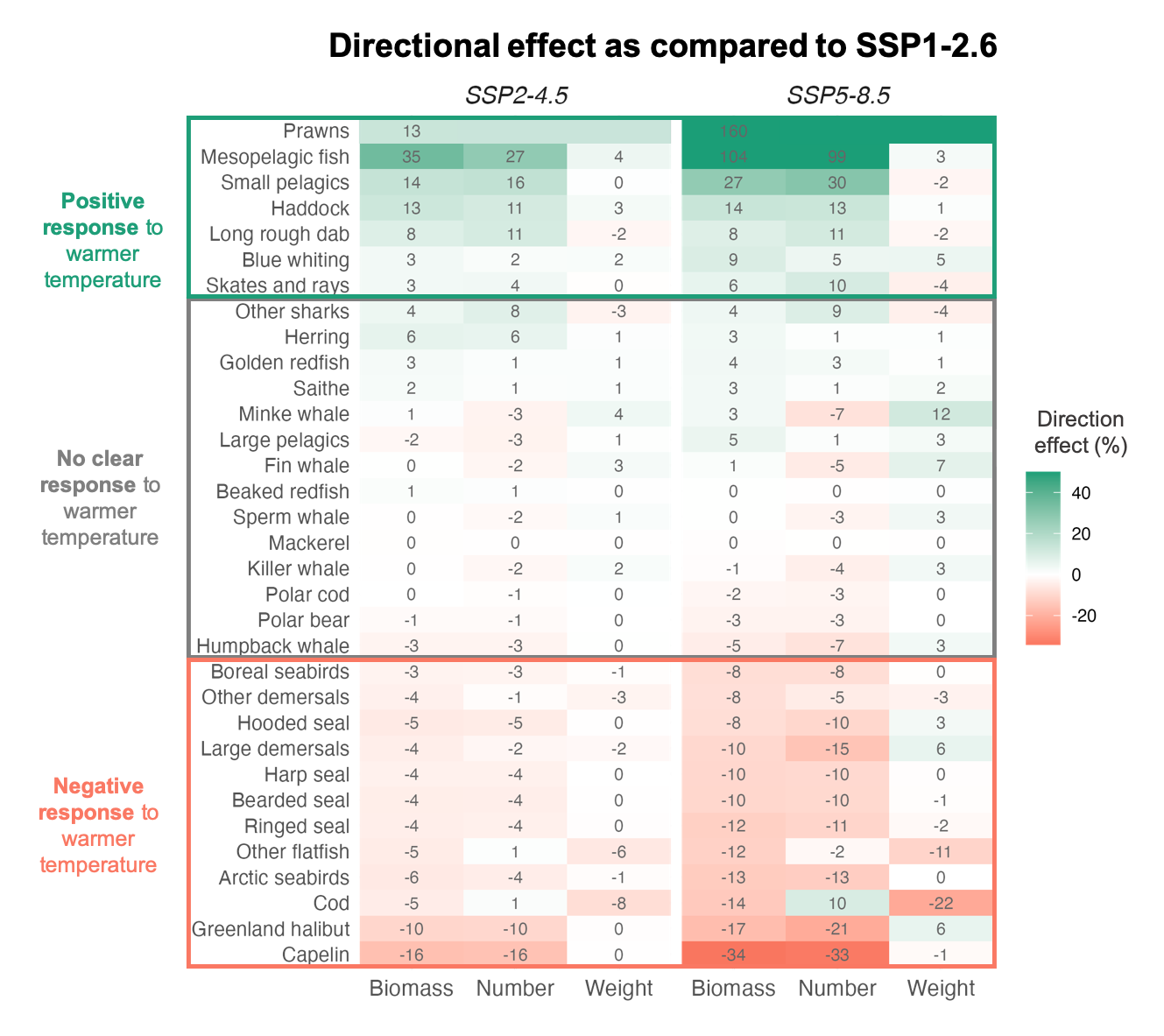
Figure 15. Species’ response to future projections, when compared to their median levels across 2090–2099 (the last 10 years of the simulation) for the SSP1-2.6 projection. The species are sorted by effect on biomass from positive response (green, increase) to negative response (red, decrease). From Nilsen et al. (2024).
Fish
It is well documented that throughout the historical record fish in the Barents Sea have been and are significantly affected by variability in their environment, particularly sea temperature, both at the single population (Hjort 1914; Ottersen et al. 2014) and community level (Fossheim et al. 2015; Ingvaldsen et al. 2021). Environmental pressures may act both directly on the fish’s physiology or indirectly through predators or prey. Impacts on individual fish may lead to responses at the population level, affecting distribution, seasonal migration patterns, and recruitment success (see recent review by Gerland et al. 2023, also Mueter et al. 2021 and Skjoldal et al. 2022)
Higher temperature, retreating sea ice, and changes in prey availability during the decade 2004–2014 affected fish especially in the northern Barents Sea. Here higher temperatures expanded suitable feeding areas for boreal/subarctic species towards the typically cold north and east and contributed to strongly increased Atlantic cod production (Kjesbu et al. 2014). In contrast, the small-sized, slow growing arctic fish species that usually dominates the region were affected negatively by a shortened ice-covered season and reduced sea ice extent through loss of spawning habitat and shelter, increased predatory pressure, reduced prey availability, and impaired growth and reproductive success (Kortsch et al., 2015; Dupont et al. 2024). The population size of the Atlantic cod is now strongly reduced and distribution retreated to “normal” areas. Still, this period of climate induced structural change over large spatial scales, leading to a borealization or ‘Atlantification’ of the European Arctic biological communities (Fossheim et al. 2015, Meredith et al. 2019, Årthun et al. 2025) may be taken as a forewarning of what will happen when more permanent climate change sets in (Fig. 16).


Figure 16. Alternative futures of Barents Sea fish with climate change (with OpenAI DALL· E 3).
It is yet difficult to distinguish between impacts of natural environmental variability, anthropogenic climate change, and other human pressures, like fishing, on the Barents Sea fish stocks. In the future, climate change will play a more dominant role, and fisheries will need to adjust to the prevailing situation (Skern-Mauritzen et al. 2016, Ottersen et al. 2025). The most significant ecological impacts from climate change on Barents Sea fish will, as many other places, be through higher sea temperatures. Temperature changes will decrease both ice cover and thickness, and affect mixed layer depth, in most cases decreasing it. This and other effects of temperature change will likely alter nutrient mixing and availability. Recent results by Sandø et al. (2024) show that future changes in mixed layer depth are a main driver for changes in NPP. Future changes in primary and secondary production will affect fish stocks, some negatively and some positively, and thus alter ecosystem dynamics in many ways. Following Sandø et al. (2024) projected increase in GSP in the Barents Sea might positively impact recruitment of boreal species like Atlantic cod and haddock and allow them to expand their distribution northwards and eastwards again.
The degree, and in some cases also the direction of change, is however, highly different not only between scenarios, which is to be expected, but more problematically also between different model approaches. While Sandø et al. (2024), as we just described, projected a positive development for Atlantic cod in a warmer ocean and Kjesbu et al. (2021) had similar results, the study by Nilsen et al. (2024) concluded differently. Their very recently published projections for the BS conducted with the NoBA Atlantis model are the first with an end-to-end model using downscaled regional physical forcing. The negative responses to warming in Atlantic cod and capelin and the unclear response of polar cod found in the NoBa Atlantis projections contrasts with the findings of Kjesbu et al. (2021) and Sandø et al. (2024). This highlights the complexity and uncertainty in present state-of-the-art long-term projections for higher trophic levels, like fish stocks.
Seabirds
Seabirds are typically at the top of the marine food web. Predicting the effects of climate variability on and through the different trophic levels is a major challenge, increasing in complexity at successively higher food web levels up to seabirds. As summarized in Ottersen et al. (2023) and reported earlier in this report for other areas, seabirds can be affected by changing climate both directly, for example if extreme weather becomes more frequent, or indirectly, through changes in their food supply. There is substantial literature pointing towards indirect effects most often being the more important of the two.
The islands around the Barents Sea (i.e., Svalbard, Franz Josef Land, and Novaya Zemlya) are the nesting places for large numbers of seabirds. Estimates of numbers are, naturally, somewhat uncertain, but likely about six million pairs from 36 seabird species breed regularly in the Barents Sea. Including also immature birds and non-breeders, the total number of seabirds in the area during spring and summer is about 20 million individuals. Note that although many species are present, 90% of the birds belong to only 5 species: Brünnich’s guillemot, little auk, Atlantic puffin, northern fulmar and black-legged kittiwake (BarentsPortal 2020). The birds feed on different pelagic ecosystem components, including zooplankton and fish. Because seabirds typically depend on rather specific prey, they may be vulnerable to changes, and function as indicators for ecosystem status (Gerland et al. 2023).
Ramirez et al. (2017) find that interannual changes in phenology, the seasonal patterns of marine productivity, may have a cascading effect on seabirds around Svalbard. In particular, they showed that increasing temporal lag between sea-ice melting, i.e. the physical process driving the annual bloom of sea ice algae, and the bloom of pelagic phytoplankton resulted in rapidly decreasing breeding performance for little auks and Brünnich’s guillemots, two of the most important and abundant species in the Barents Sea region. The timing of these two productivity pulses is considered as an essential driver of recruitment, and hence abundance. The advancement in ice breakup may result in an earlier onset of the pelagic phytoplankton bloom. This may negatively impact the seabirds both through reducing food abundance/availability and by causing a temporal mismatch between seasonal patterns in food availability and their reproductive requirements (Ramirez et al. 2017).
While the abundance of black-legged kittiwakes remains stable in most of the monitored colonies on Bjørnøya and Spitsbergen, it is declining rapidly in mainland Norway. Since 2021 they are classified as Endangered (mainland) and Near threatened (Svalbard) on the Norwegian Red List. The causes of changes in kittiwake populations are not fully known, but similar changes in populations across larger geographical areas suggest there are coinciding causal patterns. These are likely linked to changes in food availability, which are mainly related to climate, in particular changes in ocean temperature. Generally, with increasing sea temperatures and declining sea ice coverage a northward shift in Barents Sea seabird distribution is expected. This is substantiated by clear such trends for numerous species from 2009–2019 (Miljøstatus 2024).
Nilsen et al. (2024), studying future scenarios, did not focus much on seabirds, but their results for the Arctic seabirds group in the Nordic and Barents Seas as a whole point towards a rather clear negative response to warming (Fig. 15).
Marine mammals
Arctic marine mammals are large, warm-blooded and highly mobile animals that are adapted to experience significant variation in their environments. They have physiological capacities that make them quite robust to direct effects of climate change (Haug et al. 2017). Still, predicted reductions in sea ice are likely to affect seals and walruses negatively, in particular by directly reducing or removing their established breeding habitats and more indirectly by shifting the general location and timing of lower trophic level productivity (Kovacs and Lydersen 2008, Haug et al 2017).
The warming observed in the Barents Sea ca 2004–2012 had negative impact on several particularly sensitive seal species (Eriksen et al. 2021, also reported in Ottersen et al. 2023). Less ice and poorer ice quality can lead to a further decline in the populations of Greenland- and hooded seals (Eriksen et al. 2021) as well as ringed seals (Stenson et al. 2020). After the drastic changes in ice conditions that started in 2006, which could be termed a “tipping point”, the behaviour of ringed seals around Svalbard has changed a great deal. This applies to both those who migrate out and those who remain close to the coast. They now spend much more time diving and less for resting; everything indicates they are working harder to find food (Hamilton et al. 2015). Further, ice retraction from the shallow (100–350 m) shelf to the deep polar basin reduces access to bottom-associated prey species for harp seals (Haug et al. 2021) and walrus (Gerland et al. 2023). Further ahead, fertility rates, mortality rates, foraging success, and pup survival may be negatively affected for several populations of endemic Arctic marine mammals (Hamilton et al. 2015; Gerland et al. 2023).
Seals, both hooded, harp, bearded, and ringed, are projected to respond negatively to higher temperatures, thus more pronounced decline under SSP5-8.5 compared to SSP2-4.5 and especially to SSP1-2.6 is expected (Fig. 15; Nilsen et al 2024). This is directly linked to temperatures exceeding tolerated levels. However, marine mammals that depend on sea ice, especially several seal species, are also expected to decline due to habitat loss. Sea ice is important also for Polar bears in the region, still no clear response to future warming was found by Nilsen et al (2024).
The effects of warming are more uncertain for most whale species. This was stated already by Kovacs and Lydersen (2008) and clearly illustrated by Nilsen et al (2024) where all the five whale species are grouped under “No clear response” (Fig. 15). However, when projecting changes in projected core habitat for bowhead whales, Chambault et al. (2022) generally project losses over the eastern distribution region north of Svalbard to Franz Joseph Land. Between the present and 2100, their models project a significant contraction in habitat, with the most marked habitat loss suggested under scenario SSP5-8.5 (Fig. 17). The bowheads are projected to lose most of their current eastern habitat under the SSP5-8.5 scenario, while under SSP1-2.6 projections show some new habitat becoming available in the northeast (Chambault et al. 2022).
Eriksen et al. (2021) point to challenges during the early 21st century warming for beluga whales due to reduced sea ice, to bowhead whales owing to increased sea temperature and reduction in ice extent and to narwhals related to both their specialised deep-diving eating behaviour, ice dependence, and already limited distribution area.
Further, temperate whale species, including minke whales, are showing northward expansions of their ranges, which is likely to cause competition with endemic Arctic species, as well as increasing their risk of predation and diseases. Also, whale species endemic to the Arctic may face increasing competition from intensified use of (sub)Arctic habitats by seasonally migrant species, like large baleen whales (Moore and Huntington 2008, Haug et al. 2017).
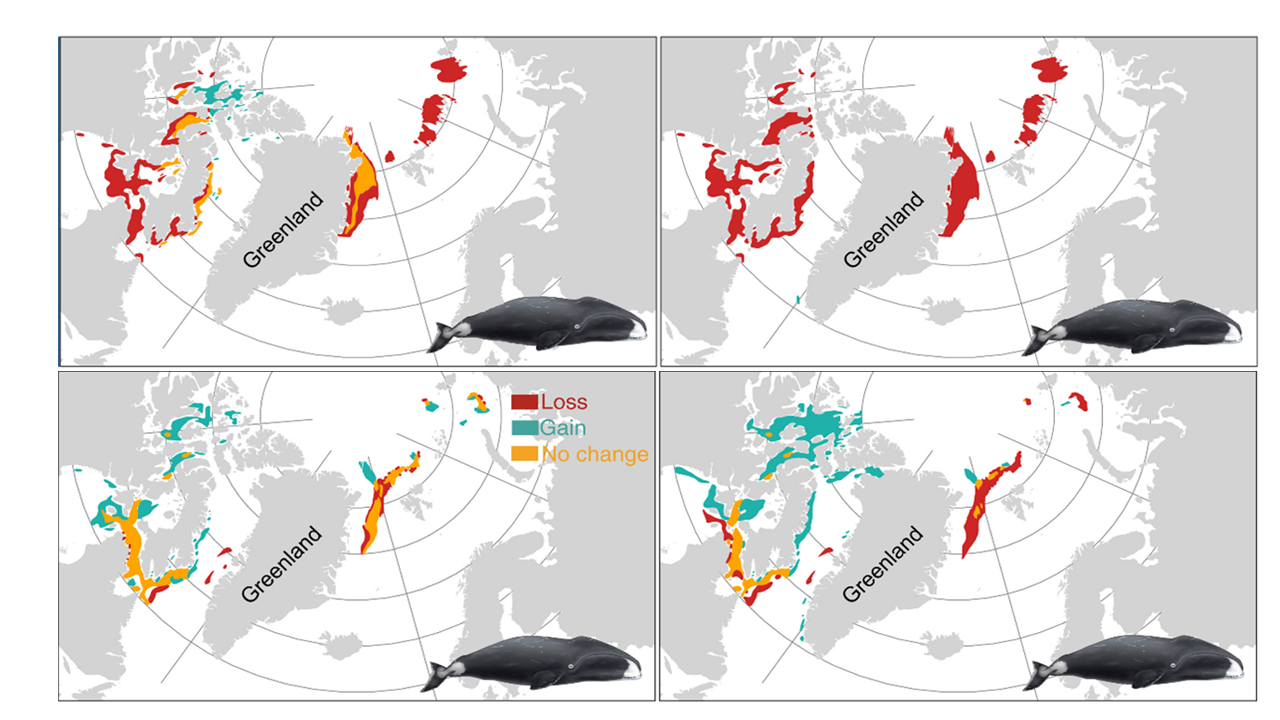
Figure 17. Changes in projected summer (upper panels) and winter (lower panels) core habitats (probabilities >0.5) by the year 2100 for Bowhead whales for the scenarios SSP1-2.6 (left) and SSP5-8.5 (right). Areas with projected losses by 2100 in red, areas expected to remain unchanged in orange, and areas with projected increases in habitat in green. Adapted from Chambault et al. 2022. Bowhead whale illustration by Uko Gorter.
Holistic ecosystem considerations
The physical changes expected in the Barents Sea are among the globally most significant. Consequently, substantial ecosystem changes are expected to arise. During the warming period from ca 2004–2012 major shifts took place, with the Atlantic fish community from the southern BS (mainly Atlantic cod and haddock) expanding towards northeast at the cost of the resident local Arctic species. A similar ecosystem change occurred for benthic populations. There was a shift northward in the amount of benthic animals and temperate benthic communities. For example, changes in benthic biodiversity was seen in the Svalbard area, again an increase in boreal and withdrawal of Arctic species. With persistent future warming such changes may become permanent in both benthic and pelagic ecosystems, for boreal fish also related to projected increase in zooplankton production (Sandø et al. 2024, Ottersen et al. 2025).
The most significant ecological impacts from climate change in the Barents Sea (as many other places) are through higher sea temperatures. Rising temperatures will reduce ice cover, thickness, and typically decrease mixed layer depth. This and other effects of temperature change will likely alter nutrient mixing and availability. In the Barents Sea, new results from the NorScen project show that change in mixed layer depth is a main driver for changes in NPP (Ottersen et al. 2025). Such changes in NPP will propagate upwards in the food web, influencing zooplankton and fish stocks. Some species and populations will be negatively affected and some positively, the ecosystem dynamics will thus be altered in many ways (Sandø et al. 2024, Nilsen et al 2024, Ottersen et al. 2025). Note that the degree, and in some cases also the direction, of change for the various species, is highly different between scenarios up to 2100.
With temperature increase, Marine heat waves (MHWs) are generally expected to become more frequent, longer lasting, and more intense. Relatively little work has been done on MHWs in the Barents sea area, but recent and ongoing research points to increasing number and severity of both surface and near-bottom events over large parts of the region (Mohamed et al 2022, Ottersen et al. 2025, S. Gonzalez, IMR work in progress). Such abrupt events may sweep through the ecosystem, causing non-linear effects that are difficult to predict or project. Although writing on the Arctic region more as a whole, the conclusions of Gou et al. (2025) are highly relevant also for the Barents Sea. They underline that the extreme temperature variations and increased stratification associated with MHWs will pose substantial challenges for Arctic ecosystems. These changes may adversely affect food webs both through direct temperature effects on physiology and indirectly by influencing nutrient supply and causing shifts in species taxonomy (Gou et al. 2025).
As high-level predators, seabird populations depend upon lower trophic levels in the ecosystem and have in high-latitude regions like the Barents Sea, evolved to fit production cycles. Climate change thus poses a particular threat to seabirds since it can disrupt the critical timing between when they need to feed and when their food sources are available (match becomes mismatch). As a result, the well-being and even survival of a seabird populations will depend heavily on the locations and timing of fish spawning, as well as the abundance of zooplankton.
Projections recently conducted with the NoBA Atlantis model are the first with an end-to-end ecosystem model using downscaled regional physical forcing (Nilsen et al. 2024). It should be noted that the negative response in cod and capelin and the unclear response of the polar cod found in the NoBa Atlantis projections by Nilsen et al. (2024) contrast with the findings in the other expert-based studies on the region (Kjesbu et al. 2021, Sandø et al. 2024). This highlights the complexity and uncertainty in long-term projections for higher trophic levels, which depend on developments in other parts of the ecosystem.
A key message from the recent extensive review by Årthun et al. (2025) is that the inter-scenario differences first become substantial late in the century. Mitigation is of great consequence for the long-term development of the ecosystems, but if most of the changes projected for 2050 are already locked in due to the inertia of the earth-ocean system, adaptation should also be an important part of management (Årthun et al. 2025). While this is written for the Barents Sea, it is likely a general conclusion and recommendation.