PART 2.
2.1 Introduction
Following the development of accurate oceanographic instruments (the Nansen bottle) in the late 1800s, a cold tongue of subarctic waters (SAW) from northeast of Iceland extending into the southern Norwegian Sea was among the first thoroughly studied oceanographic features in marine science. In their seminal book, ‘The Norwegian Sea’, Helland-Hansen and Nansen (1909) hypothesized that increased eastward extensions of this dynamic ‘Icelandic tongue’ stimulated ecosystems along the Norwegian slope and shelf. This hypothesis received little attention until recent studies by Kristiansen et al. (2016, 2019) demonstrated that the variable extent of this tongue into the waters north of the Faroe Islands regulates zooplankton communities there: a large tongue results in large biomasses of fatty and nutritious large stages of Calanus finmarchicus and Calanus hyperboreus, which can reach the Norwegian slope as well (Skagseth et al., 2022). C. finmarchicus is the critical prey item, transferring energy from primary production to higher trophic levels in the Norwegian Sea (Melle et al., 2014), and on adjacent shelves (Sundby, 2000). C. hyperboreus plays a similar role north and northeast of Iceland during spring (Gislason and Silva, 2012). The fact that these subarctic copepods vanish from the southern Norwegian Sea when the Icelandic tongue retracts westwards (Kristiansen et al., 2019), likely has significant ecological implications.
SAW circulates clockwise around the Faroe plateau, creating a complex oceanographic system that establishes the hydrographic foundation for Atlantic inflows, that pass this shelf on their journey towards higher latitudes. Confluence of lower trophic level biomass along the interface between the SAW and the overlying Atlantic water – the permanent thermo/pycnocline, feeds a large biomass of krill, mesopelagic fish and their larger predators (e.g. blue whiting, mackerel, Greenland halibut and several other fish species. This “Wall of mouths” (Hamner et al., 1988) is clearly identifiable as a Deep Scattering Layer (DSL) in acoustic data from these waters (Cisewski et al., 2021). Ascended zooplankton and other near-surface, passive, food items can potentially also be advected from the Iceland tongue towards the Faroe shelf break in a low-saline surface layer, which builds during the summer and fall (Perkins et al., 1998). A thorough discussion on the ocean-to-shelf advection is, however, beyond the scope of the present work.
Biological production on the Faroe shelf is characterized by marked peaks every 6–10 years, i.e.: 1974, 1983 (weak), 1994, 2000, 2009, 2017, and a recent weak peak in 2023–2024 (Jacobsen et al., 2019).
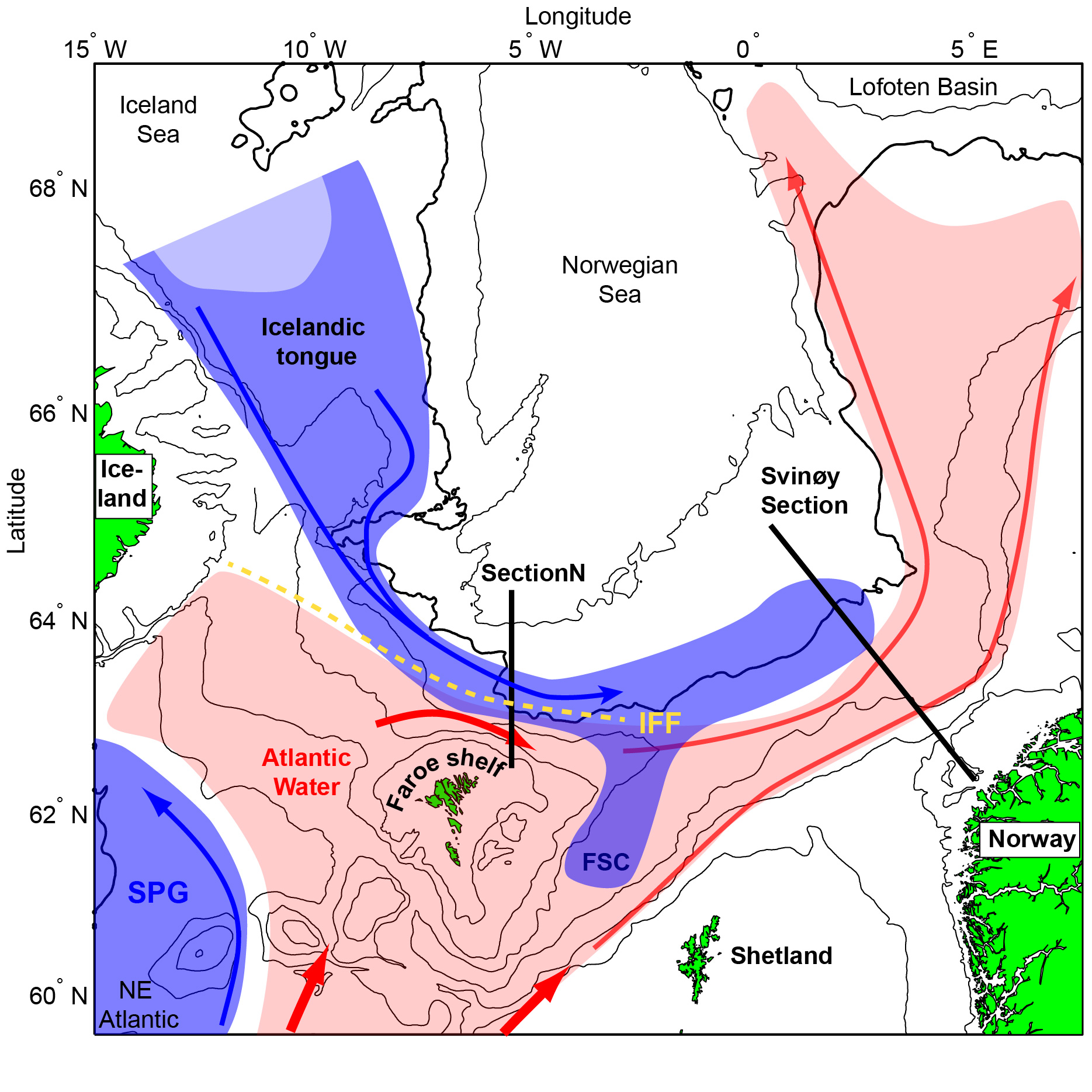
Figure 2.1 Overview over the study region. Subarctic currents and water masses are shown with blue arrows and shading, while the warmer Atlantic counterpart is shown in red colours. FSC: Faroe-Shetland Channel, IFF: Iceland-Faroe Front and SPG: Subpolar gyre.
These peaks are reflected in a so-called primary production index (PPI) (Gaard et al., 1998), and 0-group length index (Jacobsen et al., 2019) breeding success of seabirds (Olsen et al., submitted), and recruitment to commercial fish stocks (ICES, 2023). While these synchronizations are tight, the magnitude of peaks in the PPI and 0-group length index are not always reflected in recruitment. Particularly, production peaks after 2003 have not benefitted the adult cod (Gadus morhua) stock as they did before. While the Faroese fish stocks and seabird populations (biomass) exhibit rounded peaks following productive periods, they also show clear longer-term shifts and trends (ICES, 2023) (Olsen et al., submitted), which cannot be explained by the PPI or the 0-group length index.
This calls for consideration of other environmental indicators, likely with a larger-scale ‘footprint’ than just the Faroe Shelf itself. Acknowledging previously suggested links between ecological shifts in the NE Atlantic and indices like the subpolar gyre index, the North Atlantic Oscillation (NAO) index, and the Atlantic Multi-decadal Oscillation (AMO) (Hátún et al., 2009), we specifically discuss potential links to changes in SAW from the Icelandic tongue.
Guillemots (Uria aalge, hereafter guillemot) have historically constituted the largest and most exploited seabird population in the Faroe Islands (Nørrevang, 1977). A major decline in this species during the 1960s elicited concern (Olsen, 1990; Reinert, 1976) and initiated structured seabird counts in the Faroe Islands. The and the here discussed guillemot record is the longest if its kind from this shelf (Olsen et al., submitted). Cod has always been the main demersal commercial fish species, sustaining a considerable portion of the Faroese economy. This stock has therefore been closely monitored, providing extensive data material and long robust time series (ICES, 2023). Cod and guillemots share similar prey species (e.g. sandeel), and the piscivorous guillemots also prey directly on juvenile cod (Myksvoll et al., 2013), which makes the breeding performance of this seabird a useful indicator of the status and recruitment of the cod stock. Motivated by this expected ecological coupling, the economic and cultural significance, and large data availability, the total attendance of guillemots and the total cod biomass are employed as indicator records for upper trophic levels on the Faroe Shelf. With these records, we want to test the following hypothesis: the biomass of upper trophic levels on the Faroe shelf is driven by both on-shelf production and off-self food abundance.
The data material is described in Section 2.2, spatial and temporal changes in the subarctic waters are illustrated in Section 2.3.1 and updated oceanic zooplankton records are provided in Section 2.3.2. Recruitment and biomass of cod is presented in 2.3.3, and guillemot attendance is in 2.3.4. A comparison with the Faroe Bank cod is provided in 2.3.5 and the results are discussed in 2.4.