PART 1.
1.1 Introduction
1.1.1 Long term variability of marine ecosystems
There has been a long history of research on common drivers of environmental change in NE Atlantic and fluctuations in atmosperic pressure and sea temperature have received a lot of attention (e.g., Stenseth et al. 2004). One of the most notable observations was that there were long periods of herring fishery along the Norwegian coast and Iceland when temperatures were higher than normal and the conspicuous absence of herring in colder periods and curiously enough, there was an opposite pattern in the herring fishery along the Swedish west coast, i.e., a rich fishery when temperatures were low and no fishery when temperatures were high (Alheit and Hagen 2003). Although it was first proposed that this was caused by large-scale variations in migration pattern of a common herring stock in the NE Atlantic, it was later discovered that there were two stocks of herring in the areas that responded differently to temperature (Höglund 1978). Another example was the observation that the abundance of cod in the Barents Sea and in Greenland tended to show the opposite pattern (see Sundby 2000). Such large-scale differences in development of fish stocks targeted by commercial fisheries have later been attributed to variations in climate, more specifically the wind pattern and the NAO index (Drinkwater 2013). There was a large focus on the connection between NAO and fish stocks in the 1990s (Stenseth et al. 2004), but these relationships tended to break down in the 2000s. With a focus on coastal shelf ecosystems in Nordic waters, our project attempts to unravel some of the important mechanisms through which climate change drives the bottom-up productivity of marine food chains, from zooplankton to fish and seabirds.
1.1.2 Oceanography and how it affects the shelves
In our search for drivers of productivity and biogeographical changes in the NE Atlantic an obvious choice is the dynamics of the subpolar gyre (Hátún et al. 2009) . During years with strong winds and heat losses in the Labrador and Irminger Seas, intensified winter convection brings large amounts of nutrients from the deep water masses and up to the surface (Hátún et al. 2017a). This fuels a large production of phytoplankton and zooplankton within the subpolar gyre (Hátún et al., 2016; Tesdal et al., 2022) and expands the subpolar gyre eastward (Hátún et al. 2005), and fuels shelf ecosystems along boundaring the northeastern Atlantic (Hátún et al. 2022a, 2017b).
Another candidate for a driver of productivity is the strength of the East Icelandic Current (EIC). The area NE of Iceland is rich in zooplankton (ICES 2021) and advection of the zooplankton with the EIC is crucial to the production in the southern and eastern part of the Norwegian Sea. There have been marked interannual variations in the strength of the EIC where especially the years from 2004 to 2016 were characterised by low amounts Modified East Icelandic Water (MEIW) north of Faroe Islands and along the Norwegian Shelf (Hátún et al. 2022b; Kristiansen et al. 2019; Skagseth et al. 2022). A marked increase in this subarctic intermediate water mass happened subsequently in 2017 to 2021 (see Part 2).
1.1.3 Zooplankton
The copepod Calanus finmarchicus is a key secondary producer in the North Atlantic (Melle et al. 2014), and dominates the meso-zooplankton biomass in Icelandic, Faroese and Norwegian waters (Gaard and Hansen 2000; Melle et al. 2004). One of the key life history traits for this copepod species is dormancy, a strategy acquired in order to persist through seasonally adverse conditions (winter). As such, in late summer in the Northeast Atlantic, pre-adult C. finmarchicus copepodites descend to deep waters to overwinter in a resting state known as diapause (Heath et al. 2000), and the Norwegian Sea gyre and the Atlantic Subpolar Gyre are regarded central overwintering areas (Melle et al. 2014). Shortly prior to the spring bloom the animals ascend from diapause at depth to surface waters, molt into adults and begin reproduction. The egg production is to a large degree dependent on the concurrent food concentration (Stenevik et al. 2007; Head et al. 2013; Melle et al. 2014), but lipid reserves may also fuel the initial production (e.g. Madsen et al. 2008). Once in the upper layers, C. finmarchicus is advected onto adjacent shelves, where it is an important prey for larva and juvenile fish of several benthic fish species e.g. cod (Sundby 2000; Heath & Lough 2007). Furthermore, C. finmarchicus is considered a main prey for pelagic fish stocks such as herring, blue whiting and mackerel (Prokopchuk et al. 2006; Langøy et al. 2012).
The amount of zooplankton has shown similar long-term trends for large oceanic areas in the Nordic Seas. For the period 1995 to mid-2000s the plankton abundance in spring was relatively high, with fluctuations between years. Since around mid-2000s the zooplankton biomass decreased and has since been at lower levels (ICES 2024d). The period with lower zooplankton biomass coincides with higher-than-average heat content in the Norwegian Sea and reduced inflow of Arctic water into the southwestern Norwegian Sea. The arctic indicator species Calanus hyperboreus showed the same pattern, with higher abundance in the Norwegian Sea in periods with increased inflow of Arctic water (Skagseth et al., 2022). The zooplankton biomass to the south of Iceland has been shown to vary according to the strength of the SPG (Hátún et al. 2017a), and the zooplankton biomass as well as the abundance of C. finmarchicus in the southern Norwegian Sea has been linked to the EIC (Kristiansen et al., 2016; Skagseth et al. 2022). In addition, the phenology (i.e. the timing) of C. finmarchicus in the southern Norwegian Sea changes dependent on the influx of MEIW (Kristiansen et al. 2016; Kristiansen et al. 2019), which may also affect higher trophic level species. In the North Sea, the variability in abundance of C. finmarchicus has been linked to the recruitment of cod (Beaugrand et al. 2002), and for the Faroe shelf, it has recently been shown that interannual variability in advection of C. finmarchicus on to the Faroe shelf in spring affects the variability in abundance of pelagic juvenile Faroe Plateau cod (Jacobsen et al., submitted).
The zooplankton biomass and the abundance of C. finmarchicus at the south-western Norwegian shelf show a similar long-term trend as the oceanic Norwegian Sea, with lower abundance after 2010, but the signals are weaker (Dupont et al. 2017; C. Broms, not published). This may indicate that the shelf is affected by the amount of zooplankon advected from surrounding oceanic areas. However, several local factors may also play a role, as predation, and the Norwegian Coastal Current flowing along the coast in a south-north direction. For the Norwegian shelf it has been shown that the density of zooplankton in the drift route of the herring larvae dropped significantly after 2004, and their centre of gravity shifted northwards, concluding that presence of food and overlap with high food concentrations are likely important regulators of survival in herring larvae (Toresen et al. 2019).
1.1.4 Forage fish
Forage fish play a vital role in the linkage between lower and higher trophic levels of marine foodwebs (Pikitch et al. 2012; van der Kooij et al. 2008); there amongst are sandeels (Ammodytes spp.), a lipid rich schooling fish, of great dietary importance for several demersal fish stocks as well as seabirds (Eliasen 2013; Greenstreet et al. 2006; MacDonald et al. 2019).
There has been much focus on the relationship between sea temperature and the life history and ecology of sandeels in the North Sea (Réginer et al. 2018). Year class strength is estabished during early larval development and depends on the degree of temporal synchrony between hatching time and Calanus helgolandicus egg producion (Régnier et al. 2017). Rising temperatures might negatively affect the growth rate, body size and fat reserves of sandeels and, consequently, reduce both the first-winter survival of the age 0 year class (van Deurs et al. 2011) and the reproductive investment of older sandeels (Wright et al. 2017). Elevated sea temperatures also increase the metabolism of sandeels during winter when sandeels do not feed and lead to increased mortality, not only in the North Sea but also in the Faroes (Eliasen 2013) and in Selvogsbanki in South Iceland (E.S. Hansen & V. Bogason, unpublished data).
Several studies indicate that the lesser sandeel Ammodytes marinus will be vulnerable to climate change in the North Sea due to two factors. Firstly, warmer water will affect the composition of zooplankton. An increase in warm-water species and a decline in the occurrence of Calanus finmarchicus (Beaugrand et al. 2002) is expected, leading to a reduced amount of suitable zooplankton for the lesser sandeel (van Deurs et al. 2009). This could affect larval survival, growth rate, and spawning biomass of the lesser sandeel. Secondly, buried lesser sandeel will have a higher metabolism in warmer water, resulting in a reduced likelihood of winter survival and lower energy reserves for spawning (van Deurs et al. 2009; van Deurs et al. 2011; Eliasez 2013; Henriksen et al. 2021). Additionally, Johnsen et al. (2017) show that higher densities of lesser sandeel increase school formation and predator avoidance, so a general decline in the amount of sandeel will reduce school structures and natural predator avoidance. The lesser sandeel's dependence on a suitable habitat in relatively shallow water with a substrate of oxygen-rich gravel and coarse sand limits its ability to move to new areas. Therefore, it is particularly vulnerable to significant environmental changes.
In the North Sea, repeated stock assessments of the sandeel stocks have provided time series of recruitment, biomass and fishing mortality, but such data are not available for sandeels on the Nordic shelves. Although it has been acknowledged that Calanus copepods are very important prey for sandeels (Eliasen 2013) there are few studies that link sandeel abundance with oceanographic features and zooplankton advection onto the Nordic shelves (e.g., Henriksen et al. 2018). Research in the North Sea indicates that trophic mismatch of Calanus spp. and hatching time of sandeel larvae can affect recruitment of sandeels (Arnott et al. 2002; van Deurs et al. 2011; Régnier et al. 2017). Both spawning and hatching time of sandeel larvae in the North Sea have also shown correlations with bottom sea temperature (MacDonald et al. 2019).
1.1.5 Demersal fish recruitment
Fish stock assessments are routinely done for many demersal fish stocks on the Nordic shelves, e.g., cod (Gadus morhua), haddock (Melanogrammus aeglefinus) and saithe (Pollachius virens). They provide time series of recruitment, spawning stock biomass and fishing mortality. Curiously enough, all stocks seem to be special in the sense that they exhibit their own variation over time that normally is not matched by any other fish stock in the same or other areas. Reasons for this might be that the fishery itself affects the size of the spawning stock and, if the spawning stock is low, also the recruitment. The fishery might be subjected to certain management plans that affect the fishing mortality and therefore the size of the spawning stock and, probably, the recruitment. It is rare to encounter clear environmental effects on e.g. recruitment, but an obvious exception is the link between primary production and cod production on the Faroe Shelf (Steingrund & Gaard 2005) that also is seen for haddock (ICES 2023).
1.1.6 Production of seabirds
Seabirds are an ecologically diverse group of species, most of which are totally dependent on marine food sources (e.g., Croxall 1987). Their feeding ecology reflects species-specific adaptations to different foraging habitats (e.g., coastal vs. pelagic waters) and types of prey to feed their young (e.g., many small vs. few large). Parallel differences in foraging range (short vs. long trips) and foraging behaviour (e.g., surface-feeding vs. diving) also contribute to reduce inter- and intraspecific competition among individuals breeding in large colonies. The most typical seabirds are long-lived (adult survival rate of 85–95% p.a.) and reproduce slowly (clutch size of 1–3 eggs, dependig on species). This life history is favourable in environments where conditions for raising young vary markedly between years but also make the populations more sensitive to increased mortality of adults (e.g., due to disease, periods of food shortage, extreme weather events or longer-lasting effects of (other) antropogenic pressures). The current status and trends of seabirds in the NE Atlantic is of great concern (e.g., OSPAR 2024) and parallels a drastic global decrease in their population sizes for more than half a century (Paleczny et al. 2015).
The energetic requirements of seabirds peak in the chick period in mid summer (June–July) in the Nordic countries). Raising young is thus totally dependent on the availability of food within reach of the breeding site. A global study shows that the breeding success of seabirds is negatively affected when the biomass/abundance of their staple prey drops below a third of its historical maximum (Cury et al. 2011). Many seabirds feed on small fish, such as sandeels, capelin and young stages of herring and gadids, some also on large zooplankton, such as krill (e.g., Croxall 1997). The availability of these food sources is highly variable in time and space and the diet of seabirds varies accordingly, even between colonies of the same species. Sandeels are a nutrient-rich forage fish for breeding seabirds in Nordic waters. As they lie buried in the bottom sediments during winter (September to February), sandeels prefer sandy habitats, which are patchily distributed and not available in all areas. Larval and juvenile fish of other species are more important than sandeels in many areas. They are especially abundant in spring, but reach a more proftable size for seabirds in summer if their growth match in time and space with the availability of their key copepod prey, such as Calanus finmarchicus. Seabirds colonies function as important information centres where the movements of birds returning with prey guide their conspecifics towards the prey-rich areas. This reduces the time needed to locate good food patches and enables the birds to travel longer distances in their search for food, thereby reducing the risk of over-exploiting the prey sources closest to the colony (Ashmole 1963). Larger foraging areas also enable them to feed on a wider variety of prey. This may help explain cases where breeding success is better correlated with the productivity on lower trophic levels than with the abundance of their staple fish prey, or even with physical parameters such as sea temperature and salinity (e.g., Durant et al. 2003, 2006). A spatio-temporal match in trophic interactions between keystone species in the food chain is a prerequisite for high productivity of coastal ecosystems (Cushing 1990) and therefore also paramount for the success of top predators such as seabirds and demersal fish (e.g., Durant et al. 2005).
1.1.7 Research aims – the advection hypothesis
In the management of fish stocks as well as in conservation of seabirds in NE Atlantic it is imperative to understand and identify the main drivers of productivity in the shelf ecosystems. Broadly defined, the drivers can be divided into two main groups, firstly the local phytoplankton production that may regulate the production at higher trophic levels (e.g. Steingrund & Gaard 2005) and secondly, the production that originates from other places, for example by inputs of nutrients or zooplankton that are advected onto the shelves by ocean currents. In this report we will focus on the latter, not only because it may be the simpler option, but also because an analysis of oceanography, plankton, forage fish, demersal fish and seabirds apparently has not been done previously for the shelf ecosystems in NE Atlantic. In our analysis, we will study the area from South Iceland to Faroe Islands and along the Norwegian Shelf up to Lofoten Islands, i.e., we will omit coldwater ecosystems like the Barents Sea, since they might be governed by other processes. We will however consider elements of relevant processes in the North Sea, when similar data from the study area is lacking.
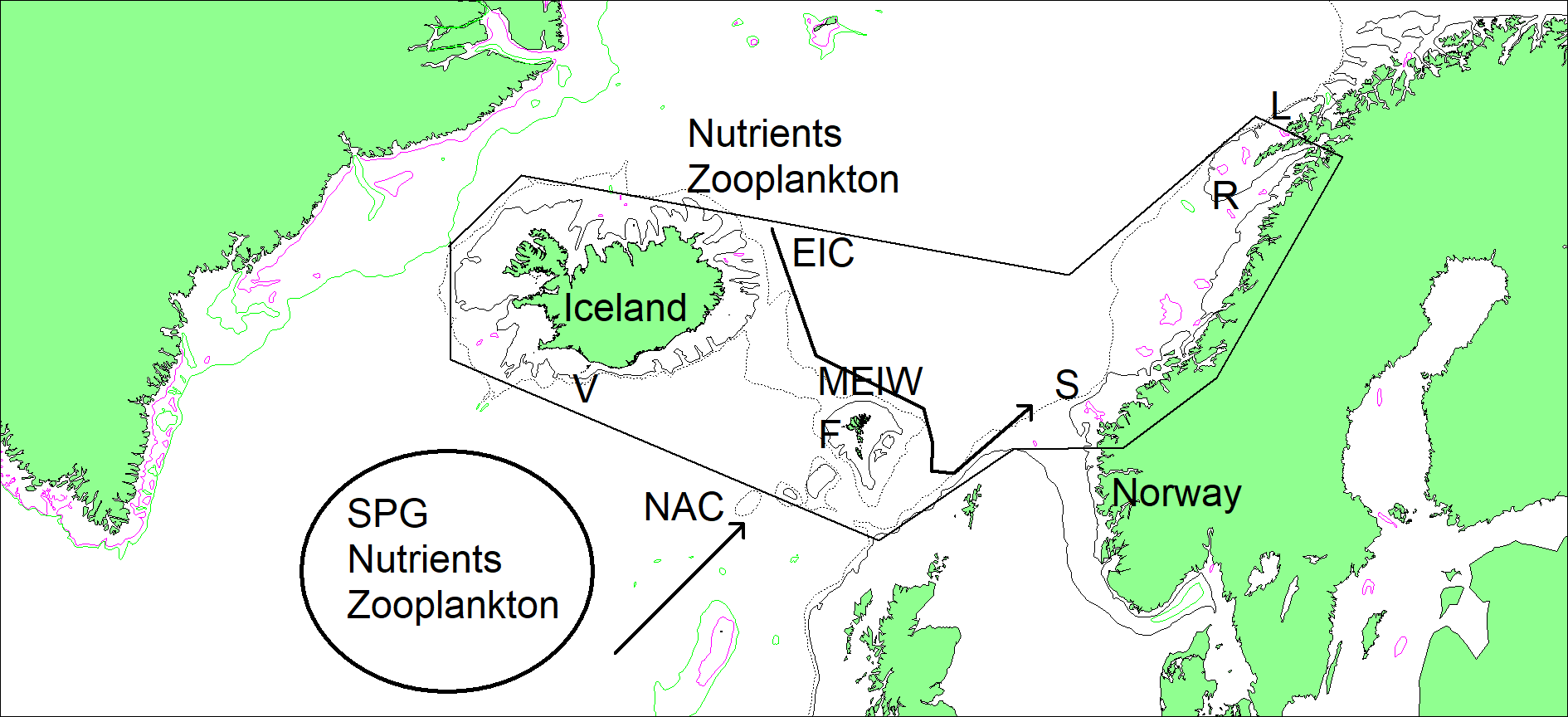
Figure 1.1 The study area (thin polygon) comprising the sea areas around Iceland, Faroe Islands (F) and along the Central Norwegian shelf up to the Lofoten Islands (L). The subpolar gyre (SPG), holding large concentrations of nutrients and zooplankton, is indicated as well as the North Atlantic Current (NAC) that flows towards the north-east. Large concentrations of nutrients and zooplankton are also found north-east of Iceland. The East Icelandic Current (EIC) transports these water masses (Modified East Icelandic Water, MEIW) towards the Faroe Plateau and onto the Norwegian Shelf, but note that not all currents are shown in this map. S denotes the location of the Svinøy hydrographical section. Large concentrations of nutrients and zooplankton are indicated. Important colonies of seabirds are found in Vestmannaeyjar (V) and Røst (R), as well as in the Faroes. For a more detailed figure, see Figure 2.1 in Part 2 of this report.
1.1.8 Materials and methods
Data on sea temperature were kindly provided by group members or from ices.dk. Data series included Faroe Mykines/Oyrargjógv annual temperature (Steingrund et al. 2024), Svinøy section temperature (ICES 2024a) and temperatures north of Iceland.
The subpolar gyre dynamics has been represented by a gyre index, based on sea surface height obtained from simulations (Hátún et al. 2005) and satellite altimetry ((Hátún & Chafik 2018; Chafik et al. 2020). These gyre indices are merged to give a continuous time series from 1960 to 2023. The recentmost data are calculated by winter (Jan)- centered annual mean altimetry values, with the mean 1994–2023 trend in the sea surface height data removed prior to the analysis.
Data on the strength of the East Icelandic Current were kindly provided by group members. This strength was quantified as the amount of MEIW north of Faroe Islands, measured as a cross-sectional area of seawater with specific temperature and salinity characteristics (1–3 °C, < 34.85) (Kristiansen et al. 2019) along standard hydrographic section N, crossing the north Faroe slope.
Data on zooplankton abundance/biomass were kindly provided by group members. They included data on zooplankton in May south of Iceland (Eyðsteinsson et al. 2021), zooplankton biomass data (large and small) on the Svinøy section (W/Norway) as well as Calanus finmarchicus and Calanus hyperboreus data north of Faroe Islands. In order to omit the shallow zooplankton data on the Norwegian Shelf, only deep stations (bottom depth larger than 250 m) were used. In order to make a common zooplankton signal for the south Iceland and southern Norwegian Sea the following series were normalised and the average taken: Icelandic zooplankton in May, Faroe Calanus finmarchicus, Faroe Calanus hyperboreus, Svinøy small zooplankton and Svinøy large zooplankton. This combined zooplankton series was compared with the zooplankton biomass index for the South Norwegian Sea that was obtained in the joint ICES coordinated ecosystem May cruise 1995–2023 (ICES 2024b).
Data on forage fish abundance/biomass were kindly provided by group members. They included sandeels in cod stomachs (frequency of occurrence) in Iceland in October and on Faroe Plateau in March and October combined.
Long-term time series of annual data on seabird performance were kindly provided by the group members and their affiliated institutions and monitoring activities. This included the longest time series data on the harvest of Atlantic puffins (Fratercula arctica, hereafter ‘puffin’) in South Iceland Vestmannaeyjar (since 1880, Hansen et al. 2021), and in Faroe islands Viðoy (since 1978) and Streymoy (since 1981). When the harvest mainly targets immature birds, these data reflect the variability in chick production of the species lagged by a few years (Hansen et al. 2021). A relative production index based on the number of chicks found in the town of Vestmannaeyjar since 1971 was also provided. In addition, there were data series on the factual breeding success for puffins in Røst, Northern Norway (since 1964), Vestmannaeyjar (since 2007) and Faroe island Mykines (since 2011), as well as for black-legged kittiwakes (Rissa tridactyla, hereafter ‘kittwake’) in Røst (since 1979) and on Skúvoy in the Faroes (since 1992). Breeding success was monitored following methods described by Walsh et al. (1995), which are an established international standard for such work. For puffins, this is based on regularly checking the contents of a selection of occupied nest burrows from the incubation period until the fate of the single offsprings (fledged/died) can be determined. The long-term metrics used were chicks fledged per egg laid (Vestmannaeyar), percentage of burrows containing a chick (Mykines) or chicks fledged per egg hatched (Røst). For kittiwakes, breeding success was determined within designated study plots based on counting the number of large, ready-to-fledge chicks per nest shown to be occupied in the incubation period. Further, site-specific descriptions of methods and materials are found in e.g., Anker-Nilssen & Aarvak (2006), Hátún et al. (2017b) and Olsen et al. (in review). By normalising the data series presented in this report, we also reduce the chance that any more subtle differences in methods (such as in techniques used to determine nests contents) or effort affect the comparison of time series across species and shelf areas.
Data on demersal fish production were obtained by ICES (ices.dk) where recruitment at age 1 was used as a measure of production. Although available we did not use stock biomass data since they may be affected by fisheries activity. We selected cod and haddock recruitment indices in Iceland (ICES 2023), Faroe Islands (Faroe Plateau) (ICES 2023) and along the Norwegian coast (Norwegian coastal cod north of 67oN) (ICES 2024c). The recruitment was set to the year of age 1 due to the indications from Faroese waters that recruitment is determined at age 1.
All of the abovementioned variables were normalised by subtracting the mean from the individual values and then dividing by the standard deviation. Averages productivity for the three shelves separately, and combined, was taken when at least two series were available. For exploratory purposes the sum of the normalised SPG and MEIW values were added together to see how a combination of the two indices compared with the average production of the trophic levels on the three Nordic shelves.
No statistics was performed although Pearson correlation coefficients were calculated to facilitate the interpretation of the results.