
- Full page image w/ text
- Table of contents
- About the contributors
- Preface
- Summary, conclusions and recommendations
- Life history and reproduction
- Impact of fishing
- Stock monitoring
- Technical regulations and closed areas
- Management units
- Effects of long-distance translocations
- Monitoring of translocations
- Understanding the loss of cleaner fish in the net pens
- Requirements for further use of cleaner fish
- Sammendrag, konklusjoner og anbefalinger
- Livshistorie og reproduksjon
- Effektene av fiske
- Bestandsovervåking
- Reguleringer og stengte områder
- Forvaltningsområder
- Langdistansetransport av fisk
- Loggføring av fisketransporter
- Behov for kunnskap om tapet av rensefisk i merdene
- Forutsetninger for en fortsatt bruk av rensefisk
- Specification of species
- 1. Introduction
- 1.1 A brief history of cleaner fish in salmonid aquaculture
- 1.2 Life history and reproduction of cleaner fish
- 1.3 Geographical distributions
- 1.4 Genetic structure
- 2. The ecological impacts of fishing and using wrasse as cleaner fish
- 2.1 Stock monitoring
- 2.2 The effects of fishing on wrasse populations
- 2.3 Wider ecological impacts of wrasse fisheries
- 2.4 Translocation and escapement from aquaculture
- 3. Management considerations for sustainable fisheries for cleaner fish
- 3.1 Overview of management in each country
- 3.2 Management units
- 3.3 Quotas
- 3.4 Size limits
- 3.5 Gear regulations
- 3.6 Bycatch regulations
- 3.7 Closed seasons
- 3.8. Closed areas
- 4. Use of wild caught and farmed cleaner fish
- 4.1. Aquaculture production of cleaner fish
- 4.2. Transport of hatchery produced Ballan wrasse and lumpfish
- 4.3. Size and transfer time for lumpfish
- 4.4. Use of cleaner fish in sea cages with salmon
- 5. Health and welfare of cleaner fish in the net pens
- 5.1 Regulatory framework
- 5.2 Animal Welfare - The cleaner fish campaign
- 5.3 Earlier assessment of mortality and welfare of cleaner fish in salmon pens
- 5.4 Operational Welfare Indicators (OWIs) and the development of better sampling regimes
- 5.5 Cataract status in lumpfish
- 5.6 Infectious diseases of wild-caught wrasses introduced into the net pens
- 5.7 Viral diseases of wrasses
- 5.8 Bacterial pathogens of wrasses
- 5.9 Parasites of wrasses
- 5.10 Infectious diseases of lumpfish
- 5.11 Bacterial diseases of lumpfish
- 5.12 Viral diseases of lumpfish
- 5.13 Lumpfish parasites
- 5.14 Transfer of pathogens between cleaner fish and salmonids?
- 5.15 Immunology and vaccine development
- 5.16 Suggested activities aiming at improving welfare and survival in captivity
- 6. Epilogue
- 7. References
- Appendix 1 - Geographical distribution of Norwegian retained catches of wild wrasse species in 2019
- Appendix 2 - Key points in the Norwegian wrasse fishery regulations for 2020. Position reporting
- About this publication
MENU
Contents
This publication is also available online in a web-accessible version at https://pub.norden.org/temanord2021-545.
About the contributors
NORWAY
Stein Mortensen is a senior scientist at the Institute of Marine Research (IMR), Bergen, Norway. His research fields are diseases of marine organisms, mainly bivalves, disease transmission and research on oysters. He has been leading several projects funded by The Nordic Council of Ministers. Project leader and main editor of the report.
Kim Tallaksen Halvorsen is scientist at IMR. His research is focused on understanding the interplay between life history, reproduction, population dynamics and fisheries in wrasses. He leads IMRs project on developing new computer vision tools for camera-based monitoring of wrasses and other coastal species.
Anne Berit Skiftesvik is a principal research scientist at IMR, Austevoll, Norway. She has been working with wrasses since mid-1990’s, both in an aquaculture (farming and use) and wild stock setting, and she heads IMR’s research and management advice on wrasses.
Caroline Durif is a principal research scientist at IMR, Austevoll. Her research deals mainly with life history strategies, migration of diadromous fish and other coastal species. She studies wild lumpfish mainly for stock assessment and advice for commercial fisheries.
Lars Helge Stien is a scientist at IMR, Matre research station, working with fish welfare in aquaculture. He has worked both with salmon and cleaner fish welfare in sea cages, and welfare effects from various preventive and control strategies against lice.
Eeva Jansson is a postdoctoral researcher at the Institute of Marine Research, Bergen, Norway. She is a population geneticist studying wild cleaner fish populations. She contributed to sections dealing with population genetics.
María Quintela is a scientist at IMR, Bergen working on population genetic studies in different fish and sea mammal species.
Nina Sandlund is a scientist at IMR, Bergen, Norway working with fish diseases. Her main research topics have been bacterial infections in various species of both wild and farmed fish.
Kjell Nedreaas is a principal scientist at the Institute of Marine Research, Norway. His research interests include fisheries dynamics and ecosystem management.
Terje Jørgensen is a principal scientist at the Institute of Marine Research, Norway. His main field of research is responsible harvesting and gear technology. He has mainly contributed to the studies on harvesting and gear selectivity.
Hans Hagen Stockhausen works as scientist at the Institute of Marine Research, Norway. With a background in geophysics his research interests among others lie in migration processes of fish and possible links to the earth's magnetic field.
Kjetil Korsnes is an associate professor II at Nord university, Bodø, Norway. His field of science are diseases of marine organisms, diagnostics, pathogen transmission and host-pathogen interactions. Contribution to the section with lumpfish.
Inger Fyllingen is a senior advisor in the head office's section for fish health and fish welfare at the Norwegian Food Safety Authority. Her main topic is fish welfare, according to the Norwegian Animal Welfare Act, including regulations of fish welfare.
Hulda Bysheim is a veterinarian and senior adviser at the Norwegian Food Safety Authority. She works with regulating the fish farming industry according to the Norwegian regulations regarding fish health and welfare.
Patrick Reynolds is a senior scientist at Gildeskål Forskningsstasjon AS (Gifas), Inndyr, Norway. Since 2010, principally involved in lumpfish research.
SWEDEN
Ellika Faust is a PhD student at Tjärnö Marine Laboratory, Gothenburg University. Her research is focused on population genetics in marine organisms, especially cleaner fish. She has contributed to sections regarding population genetics, management units and quotas, translocation and escapement, and sections concerning Swedish fishery and management.
Håkan Wennhage is an associate professor working with research and advice at the department of aquatic resources, SLU, Sweden. His research includes the importance of coastal habitats for commercial species and more broadly ecosystem effects of fisheries.
Carl André is a professor in marine evolutionary biology at Tjärnö Marine Laboratory, Göteborg University. He investigates stock structure and local adaptation in marine fish and shellfish and apply this knowledge in management.
DENMARK
Peter Rask Møller is an associate professor and curator of fishes, herpetology and mammals at the Natural History Museum of Denmark, University of Copenhagen. Since September 2020, also Associate Professor II at the Norwegian school of Fisheries, UiT Norwegian Arctic University. Since 2006 he has been responsible for The National Danish Fish Atlas and since 2019 of a EHFF project about wrasses in Denmark. Has contributed to the sections about Denmark.
Henrik Carl is a scientific employee at the National History Museum of Denmark, University of Copenhagen. Since 2006 he has been in charge of The National Danish Fish Atlas and since 2019 has also worked on a EHFF project about wrasses in Denmark. Has contributed to the sections about Denmark.
Jacob Linnemann Rønfeldt is a biologist and project employee at the National History Museum of Denmark, University of Copenhagen. Since 2019 he has worked on a EHFF project about wrasses in Denmark. Has contributed to the sections about Denmark.
ICELAND
Albert Kjartan Dagbjartson Imsland heads the Icelandic office and is the research manager at Akvaplan-niva, Tromsø and associate professor II at University of Bergen, Norway. His field of science are central physiological and endocrinological processes related to normal growth, development and reproduction of important aquaculture species. Also, population genetics of aquaculture species and possible utilization of genetic differences for aquaculture production. Contribution to the section on lumpfish.
UK
Lauren Henly is a PhD student at the University of Exeter, UK. Her research investigates the state of the fishery for wrasses on the south coast of the UK, its social and environmental impact, and options for sustainable management. She is using approaches from multiple disciplines to monitor the English fisheries, assess connectivity of populations of wrasses and trophic ecology, and investigate the importance of wrasses to the ecosystem and human stakeholders. She has contributed to the sections on the UK.
Preface
Parasitic Salmon lice cause great economic losses in salmonid aquaculture and has a negative effect on wild salmon and trout populations. The use of cleaner fish to de-louse farmed salmon and rainbow trout has rapidly increased in Norway during the last decade, coinciding with the weakened efficiency of various pharmaceutical treatments.
Among the wrasses, relevant cleaner fish are the goldsinny wrasse, the corkwing wrasse and the ballan wrasse. Two other wrasse species - the rock cook and the cuckoo wrasse - are caught as by-catch but rarely used as cleaner fish; and are therefore not the focus of this report. The use of lumpfish has increased in recent years and farmed lumpfish has surpassed wrasse in numbers. Wild lumpfish are being fished to sustain broodstock, and issues regarding translocation and escape can therefore be relevant also for this species.
This project, and report, was initiated by researchers at the Institute of Marine Research (IMR) and builds upon several years of scientific work linked to the biology, fisheries and use of wild wrasse and lumpfish. Interactions with colleagues from Denmark, Sweden, Iceland, and the United Kingdom highlighted critical knowledge gaps which must be addressed to achieve sustainable fisheries of cleaner fish with minimal negative environmental impacts. Funding from the Nordic Council of Ministers enabled us to establish the present network of scientists and managers. Key topics were identified and discussed in meetings and workshops during the project period.
The report provides a state-of-knowledge on the biology of Nordic cleaner fish, challenges regarding environmental impacts of fishing, translocation and how management have dealt with the fisheries in the Nordic countries. The development and challenges related to the use and welfare of cleaner fish in captivity are also presented and discussed. By our shared knowledge and experiences, we provide specific advice and recommendation on key management considerations. Thus, the report may also be relevant for researchers and managers in the United Kingdom and Ireland, where cleaner fish are fished and used as well.
We have entered the United Nations’ Decade of Ocean Science for Sustainable Development. The objective for this project was to establish a common platform of knowledge that would aid as a basis for developing a sustainable fishery and use of cleaner fish. The fisheries for wrasse are relatively strictly regulated in Norway and Sweden and is based on scientific advice. However, the management strategies and survey methods differ between Norway, Sweden, and UK, and we believe that the information presented in this report can be helpful for improving the assessment and regulations in each country, to minimize the risk of negative, long-term impact on the target species and the coastal ecosystems.
The use of cleaner fish creates many potential risks and challenges to sustainability. Human activities should not lead to detrimental, long lasting, or permanent changes in the resources / populations, like:
- Establishment and spreading of new diseases
- Permanent changes in population genetics
- Depletion of stocks to a level from which they will not recover
- Irreversible, indirect impacts on other parts of ecosystems due to point 1-3.
The long-distance transportation of cleaner fish comes with the risk of introducing new diseases to captive and wild fish population in the recipient areas. Another challenge is the escapement of translocated cleaner fish, which has been shown to affect the genetic structure of corkwing wrasse at the northern range end. To be able to evaluate whether the use of non-local cleaner fish can be sustainable, there is a need for more research on the risk and potential consequences, as well as dedicated work by the industry to reduce the probability that cleaner fish escape from the farms.
There are considerable challenges with the animal welfare of cleaner fish. The mortality is much higher than for farmed salmon and the welfare and disease situation of cleaner fish has been debated and questioned by NGOs and public media in recent years. The Norwegian Food Safety Authorities has stated that if the welfare is not improved and the effect of cleaner fish better documented, it can be necessary to reduce or even terminate of the use of cleaner fish. This will of course have implications for the Swedish and potential future Danish fishery.
The future use of cleaner fish depends on a proactive and transparent industry that is dedicated to rapid improvement of the current situation for cleaner fish in captivity. Practical solutions and inventions to these challenges should be well documented and scientifically evaluated, which require close collaboration with the relevant research institutes and managers. The public debate on the ethical dilemmas and welfare challenges for cleaner fish should be a catalyst for change and improvement.
Bergen, 13 October 2021.
Stein Mortensen
Summary, conclusions and recommendations
Parasitic salmon lice cause great economic losses in the aquaculture of Atlantic salmon and rainbow trout. It also has a significant impact on wild populations of salmonids, particularly sea trout in areas with aquaculture activity. Several pharmaceuticals have been used for treatment of salmon lice infestations, but over time the lice have developed resistance to these treatments, and there is a growing concern regarding the environmental impact of chemical and pharmaceutical treatments on non-target organisms. Consequently, there has been a strong incentive to find alternative methods for de-lousing. The use of cleaner fish, which pick lice from the skin of infested salmonids, has become an important tool to fight lice.
Lumpfish and several species of wrasses are used as cleaner fish. There are extensive fisheries for wrasses in the UK, along the Swedish west coast and in Norway. In addition, there is aquaculture of ballan wrasse and lumpfish in Norway, and lumpfish in the UK and Iceland. The fisheries and use of cleaner fish have evolved relatively fast, whereas scientific data collection, as well as the development of a regulatory framework, has lagged behind. Challenges linked to poor welfare, transmission of diseases and changes in the genetic structure of local populations as a result of translocations have raised questions regarding the sustainability of the current cleaner fish practice.
Both ballan wrasse and lumpfish are relatively new as farmed species, and as new species in aquaculture, there are knowledge gaps that must be filled. Research on different aspects for each of the two species has recently increased to address problems and prepare the fish for a life as cleaner fish in sea cages. There is also an increase in research that aims to increase survival and welfare of cleaner fish. There is still a way to go, but information from farmers who have good results show that it should be possible to achieve improvement. A wide range of research projects and studies have been initiated in Scandinavia and the UK over the last decade. In order to facilitate a more rapid spread and implementation of good management solutions that are based on knowledge of the species, it is therefore important that scientists and regulating authorities in different institutions and countries have a common knowledge-base and are collaborating efficiently. This report presents the state-of-knowledge on the biology of cleaner fish, the challenges regarding environmental impacts of fishing, translocation and how management have dealt with the fisheries in the Nordic countries. The development and challenges related to the use and welfare of cleaner fish in captivity are presented and discussed. Based on our shared expert knowledge, we hopefully provide management recommendations on how the fisheries and use of cleaner fish can become more sustainable. A sustainable fishery should allow the target species to replenish and have no long-term or permanent negative changes on ecosystem diversity, function, or productivity. A sustainable practice should minimize the harm and death of cleaner fish, as well as escapement and the associated risks; including transfer of new diseases to new species and areas, and irreversible genetic changes in the wild populations that reduce fitness or adaptability.
The report also identifies data gaps that may be filled with future, hopefully collaborative, research or monitoring activities. Although there is an active collaboration between Swedish and Norwegian cleaner fish researchers there is still a considerable potential for improvement through exchange of knowledge and experiences between scientist and managers in Norway, Sweden and the UK, as well as with those in other countries with developing or emerging fisheries for wrasses, like Denmark.
Life history and reproduction
Wrasses show an extraordinary variation in life history traits and reproductive strategies. This complicates sustainable management of a multispecies fishery. Better knowledge on life history variability between and within species is therefore essential for understanding how the different species are affected by fishing. Data on growth, life span, size/age at maturity and sex change is lacking for ballan wrasse in most areas where it is fished. Better knowledge on the importance of nesting males in reproduction (ballan and corkwing wrasse) is needed, as males appear to be disproportionally targeted due to their larger size. Routine, standardized sampling programmes of fish scales and/or otoliths should be established in key areas, ideally in conjunction with sampling for genetic structure, as life history traits may differ among genetically isolated populations. Protocols for sampling should be similar across countries (gear type, sampling time, sexing, ageing etc.), and data should be shared to facilitate a broader understanding of the underlying mechanisms for life history variability.
Impact of fishing
Case studies have shown that the effects of fishing on local populations can be variable, ranging from no discernible effect to considerably affected. The fishery is species-, size- and sex-selective, which can lead to unbalanced harvesting and reduced reproductive potential. Corkwing and ballan wrasses are the species with the highest depth overlap with the fishery and may therefore be most vulnerable to depletions. Some marine reserves have been shown to have higher abundance and larger wrasses compared to nearby areas open for fishing, while other studies have found no such differences. This probably reflects a highly heterogenous distribution of fishing effort between and within countries and fishing regions. Further, wrasses show very high residency, and there is limited interchange of juveniles and adults between nearby populations. Consequently, it might take a long time for overfished areas to be recolonised from adjacent sub-populations. The impacts of fishing can therefore be local and difficult to detect and manage. However, undersized wrasses will still be there and form the basis for wrasse in those area. If fisheries of wrasses change species composition and/or size structure in communities of wrasses, this may in turn affect functional ecology and trophic interactions. Wrasses feed on crustaceans, molluscs, gastropods, hydrozoans and polychaetes and are important prey for larger piscivorous species. However, there are few published studies on the ecological links between wrasses and other ecosystem components that go beyond correlations and descriptions. In Southern Scandinavia, the abundance of wrasses has shown an increasing trend over the last 30 years, coinciding with the decline of large piscivores. In addition, climate change, which can result in warmer seawater, will have a positive effect on wrasse stocks. More research is required to obtain a deeper understanding of the trophic role and interactions of the wrasses, as well as how the fisheries are affecting them.
Stock monitoring
The monitoring of abundance and size structure of exploited stocks is important for sustainable fisheries management. We recommend that both fishery-dependent and fishery-independent surveys are carried out to monitor the changes in the wrasse stocks. A survey aiming to produce a stock indicator for wrasses should use standardized gears and utilize data loggers to obtain associated environmental variables, such as temperature, salinity, and depth, that are known to affect catch rates of wrasse. Length data also has high value and gives the possibility to use length-based stock assessment models, which may be appropriate for the wrasse species. Fishery-independent surveys, conducted within and outside marine protected areas are effective for assessing the impact of exploitation, given the high site fidelity of these species. Replicated study designs and collection of data on the fishing intensity in control sites (past and current) would be advantageous. Comparative studies including several sampling methods are recommended to quantify bias of the different survey methods. Video surveys combined with computer vision for automated counting can be a promising alternative to catch-based surveys or snorkelling and should be further developed.
Technical regulations and closed areas
Technical measures regulate how, where and when fishers may fish, and varies considerably between countries. It is recommended that the full range of technical measures for wrasse management are exploited. Minimum size limits should reflect each species size at maturity, and maximum size limits should be used to protect males in those species or populations with strong sexual dimorphism (ballan wrasse, corkwing wrasse). Gears should be designed to reduce bycatch and catch of large ballan wrasse and to allow small wrasse to escape. Bycatch and smaller wrasse should be immediately and carefully released at the site of capture. Bycatch of other species constitute 10-20 % of the catch (in numbers) in the Norwegian fishery. Similar assessment of bycatch should be done in other countries. Studies investigating post-release survival of key bycatch species under different release scenarios is needed. Fishing during the spawning period should be avoided. The onset and duration of the spawning period may vary geographically and between years, so it is advised to conduct a weekly scientific survey to assess this or to use a conservative start date. Closed areas are efficient in preserving natural species composition and size structure, viewed as a positive supplement to other management measures and have particularly high value for scientific surveys monitoring the impact of fishing.
Management units
The natural population structure should be considered when defining management units. It is important that management units are based on meaningful biological entities, such as genetically differentiated populations. These units are likely species-specific so that generalizations are hard to make. Larval and egg dispersal should be taken into consideration when defining appropriate management units for the different species. When this knowledge is available, it should be effectively communicated to the relevant management authorities for a reconsideration of the spatial distribution of fishing effort and regulations.
Effects of long-distance translocations
Most wrasses used in Norwegian salmon farms have been caught locally, but around 25 % are imported from Southeastern Norway and Southwestern Sweden. Translocation of cleaner fish between distant populations can affect native populations through disease transfer and result in irreversible genetic changes if they escape. Repetitive transportations increase the risk of introduction and spread of pathogens. The introduction of non-native species in mixture with pathogens may promote host-switching and thereby the emergence of new and known diseases in new hosts. The current farm practice with its open design increases the risk of transmission between farmed and wild fish by enabling pathogens to disperse to the surroundings. In addition, the high density of hosts in a farm situation increases disease prevalence.
Recent genetic studies show that cleaner fish escape and reproduce with local populations in the northern range, which can result in genetic changes and reduced fitness in the local populations of wrasses. In particular, relocation of wrasse to the edge of each species distribution, where local fish are sparse, should be avoided since escaped fish can have an impact on here. Currently, it is permitted to import wrasse from Sweden during the spawning period – which probably increase the likelihood of genetic introgression. Thus, we recommend harmonizing the opening dates of the fisheries in Norway and Sweden to reduce this risk. Relocation of wrasse to the edge of each species distribution, where local fish are sparse, should be avoided.
Monitoring of translocations
Norwegian transporters do not have to log and report the source, or the destination of wild-caught cleaner fish caught in Norway. Given that the strongest discontinuity in genetic structure is within Norway, on the southwest coast between Skagerrak and the coastline North of Jæren, it is important to know the destination of wrasses caught along the Skagerrak coastline. In order to assess the hazards linked to translocations, knowledge on the source and destination of all wild caught cleaner fish is a prerequisite. A possible solution may be that the farmers must report the origin of the fish they release in the net pens.
Understanding the loss of cleaner fish in the net pens
There is a high loss of cleaner fish in the net pens. The proportion of loss of cleaner fish due to escapes or mortalities is poorly known. A recent study by the Norwegian Food Safety Authorities (NFSA) revealed that the farmers registered an average mortality of around 40%, but that they believed the actual mortality was closer to 60%. This discrepancy illustrates that the current registration and reporting systems of mortality in cleaner fish are insufficient. This leads to poor data quality and makes it difficult to identify the causes of the high mortality and the actual proportions of escaped and dead fish.
The farmers reported to the NFSA that they consider de-lousing, diseases, and handling as the main causes of mortality. There are presently no adequate systems for sufficient monitoring of disease outbreaks among cleaner fish. Mortalities and escapes of cleaner fish are often not detected until long after the losses occurred. New systems for real time surveillance of cleaner fish, for example based on machine vision and artificial intelligence, can contribute to better documentation of mortality, fish health and welfare.
There should therefore be developed systems for more accurate registration of mortality and disease for cleaner fish in sea cages. In addition, today’s reporting system of fish biomass and mortality to the government is outdated, making the data difficult to analyse. It is thus urgent for both the farmers and the authorities that new systems for registration of cleaner fish mortality in salmon and rainbow trout sea cages are developed.
Requirements for further use of cleaner fish
The Norwegian Food Safety Authority (NFSA) has stated that the use of cleaner fish cannot continue unless there is a significant improvement in welfare as well as a better documentation of cleaner fish as a de-lousing method. The Norwegian Council for Animal Ethics has recently come to a similar, but slightly more restrictive conclusion, in a recent recommendation the council points to a lack of knowledge, both with regards to actual mortality, the effect of cleaner fish to remove lice and best practice to secure the cleaner fish species requirements for environment and handling conditions. They recommend limitations in the use of cleaner fish until better welfare and effect can be documented. The need for improvement in the welfare of cleaner fish was thereafter laid down in the new Norwegian, governmental strategy for the aquaculture, which states that “... If the industry in the coming years cannot document in a satisfactory way that the cleaner fish can live good lives in the net cages and contribute significantly to fight salmon louse, the government see it as a natural consequence that the use of cleaner fish must be terminated”(own translation).
To fulfil the requirements signalized by the authorities, it is necessary to show that it is possible to give the different cleaner fish species a good life in the sea cages, improve best practice manuals and guidelines for cleaner fish, and obtain documentation of how efficient the different species are, and if the use of cleaner fish contributes significantly to reduce the number of salmon lice.
It should be clarified whether the high mortality that the farmers report is a result of factors that may be improved through better management and farming practices, or if the cleaner fish species are simply not able to adapt to farming conditions (are not domesticated).
Sammendrag, konklusjoner og anbefalinger
Den parasittiske lakselusen er årsak til store økonomiske tap i oppdrett av laks og regnbueørret. Lakselusen er skadelig for ville bestander av laksefisk, særlig for sjøørret i områder med oppdrettsaktivitet. En rekke kjemiske lusemidler har vært brukt for å fjerne lakselus. Dette har gjort at lakselusen har utviklet motstandskraft mot behandlingene, og det er økende bekymring knyttet til den effekten lusemidlene kan ha på miljøet, og særlig andre organismer i nærheten av oppdrettsanleggene. Det har derfor vært et påtrykk for finne alternative metoder for lusebehandling. En rekke alternative metoder er utviklet, og bruk av rensefisk – fisk som plukker lakselus fra laksens hud – er blitt et viktig verktøy i kampen mot lakselusen.
Flere arter av leppefisk og rognkjeks brukes som rensefisk. Det har vokst frem et betydelig fiskeri av leppefisk i Storbritannia, langs den svenske vestkysten og i Norge. I tillegg er det etablert oppdrett av berggylt og rognkjeks i Norge og rognkjeks i Storbritannia og på Island. Fisket etter leppefisk og bruken av rensefisk har utviklet seg raskt, og verken innsamling av vitenskapelige data eller utviklingen av et regelverk rundt denne næringen har holdt tritt med utviklingen. På bakgrunn av utfordringer knyttet til dårlig fiskevelferd, sykdom, smittespredning og genetiske endringer i bestandene blir det også stilt spørsmål om dagens bruk av rensefisk er bærekraftig.
Både berggylt og rognkjeks er relativt nye som oppdrettsarter, og som nye arter innen havbruk er det kunnskapshull som må fylles. Forskning på forskjellige aspekter for hver av de to artene har økt for å håndtere problemer og forberede fisken til et liv som rensefisk i merdene. Det er økt forskning som har som mål å øke overlevelsen og velferden til rensefisken i laksemerdene. Det er fremdeles en vei å gå, men informasjon fra de oppdretterne som har gode resultater, peker på at det kan være mulig å få til en bedring. En rekke studier og forskningsprosjekter er blitt gjennomført de siste årene, både i Norden og i Storbritannia. For å sikre en hurtig kunnskapsspredning og implementering av gode forvaltningsmodeller er det viktig at forskere og forvaltere i forskjellige institusjoner og land har en felles kunnskapsbase, kommuniserer og samarbeider på en hensiktsmessig måte. Denne rapporten presenterer kunnskapsstatus på rensefiskens biologi, utfordringer knyttet til miljøeffekter av fisket og flyttinger av fisk, og ser på hvordan forvaltningen av fiskeriet har utviklet seg i de nordiske landene. Bruken av rensefisk og utfordringer knyttet til velferd er presentert og diskutert. Basert på forfatternes kunnskap gir vi råd og anbefalinger om hvordan fiske og bruk av rensefisk kan bli mer bærekraftig. Et bærekraftig fiskeri må bygges på at de artene som fiskes klarer å opprettholde bestandene og ikke føre til langvarige eller permanente negative endringer på økosystemenes diversitet, funksjon eller produktivitet. En bærekraftig praksis forårsake minimal sykdom og dødelighet, unngå rømminger og faremomenter knyttet til dette, som overføring av sykdom og permanente endringer i ville bestander som forårsaker redusert fitness eller tilpasning.
Et mål med rapporten er også å identifisere kunnskapshull som vi forhåpentligvis kan tette ved å samarbeide om forskning og overvåkingsaktiviteter. Selv om det har vært et samarbeid mellom svenske og norske leppefiskforskere er det fremdeles muligheter for forbedring gjennom økt kunnskapsutveksling og deling av erfaringer mellom svenske, norske og britiske fagfolk, samt med fagmiljø i de landene som etablerer og utvikler et leppefisk-fiskeri (som i Danmark).
Livshistorie og reproduksjon
Leppefiskene har en stor variasjon i livshistorie og reproduksjonsstrategier Dette kompliserer en bærekraftig forvaltning, siden fisket omfatter flere arter. For å kunne forstå hvordan de ulike artene blir påvirket av fisket trenger vi bedre kunnskap om de ulike artenes biologi, og variasjoner innen artene. Særlig for berggylt mangler vi mye grunnleggende data om vekst, maksimal alder, størrelse/alder ved kjønnsmodning og kjønnsskifte. Det trengs også mer data om rollen til redebyggende hannfisk (hos berggylt og grønngylt), siden hannfiskene er større og mer fangstbare enn hunnene ved samme alder. Prøvetaking av fisk i nøkkelområder (skjell, otolitter, prøver til genetiske studier osv) bør standardiseres. Dette kan legge grunnlaget for å studere forskjeller i livshistorien til genetisk isolerte bestander av fisk fra ulike områder i hele Norden og beskrive de underliggende mekanismene for variasjon. Prøvetakingsprotokoller bør altså være lik i ulike land (redskapstyper, prøvetakingstidspunkt, kjønnsbestemmelse, aldersbestemmelse osv) og data bør deles.
Effektene av fiske
Vi har begrenset kunnskap om effektene av fisket. Studier har vist et fiskeripåvirkningen varierer betydelig fra område til område, og spenner fra ingen målbar effekt til betydelig fiskedødelighet. Fisket er arts-, størrelses- og kjønnsselektivt. Dette kan resultere i ubalansert uttak og innvirke på bestandenes reproduksjon. Grønngylt og berggylt er de leppefiskartene som i størst grad har en utbredelse som overlapper med områdene som fiskes, og kan derfor være sårbare for lokal nedfisking. Noen studier har vist at marine verneområder har høyere tetthet og større fisk enn omliggende områder som er åpne for fiske, mens det i andre områder ikke har vært funnet slike forskjeller. Dette reflekterer at fiskertrykket er ujevnt fordelt mellom og innad ulike land og regioner. Hvis fisket fører til endringer i artssammensetningen og/eller bestandsstruktur kan dette påvirke økologi og trofiske interaksjoner. Leppefisk beiter på krepsdyr, bløtdyr, børstemark og andre små virvelløse dyr. De er selv viktige byttedyr for større rovfisk. I den sydlige delen av Skandinavia ser mengden av leppefisk ut til å ha økt de siste 30 årene. Samtidig har bestandene av store rovfisk avtatt. Det finnes få publiserte studier av koblingene mellom leppefisk og andre arter, ut over det rent beskrivende. Det trengs derfor mer forskning for å få en grunnleggende forståelse av leppefiskenes rolle og samspill med andre arter, samt hvordan samspillet påvirkes av fisket.
Bestandsovervåking
Overvåking som gir data om utbredelse og størrelsen av de leppefiskbestandene det fiskes på er nødvendig for å oppnå et bærekraftig fiske. Vi anbefaler at det gjøres både fiskeriavhengig og -uavhengig overvåking for å beskrive endringer i leppefiskbestandene. I en overvåking som skal etablere en bestandsindikator for leppefisk må vi standardisere fangstredskapene og bruke dataloggere for å få miljødata som temperatur, salinitet og dyp. Også lengdedata gir verdifull informasjon, og muligheten for en lengdebestemt bestandsestimering av leppefisk. Fiskeri-uavhengig overvåking, i og utenfor marine verneområder er effektiv for å måle effekten av fisket, siden disse artene er stasjonære.
Gjentatte studier og innsamling av data på fiskeintensitet i kontrollområder (før vs. etter) vil være fordelaktig. Sammenliknende studier som inkluderer ulike metoder er anbefalt for å beregne usikkerhet, variabilitet og muliggjøre sammenlikning av data fra ulike målinger. Video-overvåking, kombinert med dataanalyse for automatisk telling kan bli et alternativ til fangstbaserte metoder eller snorkling og bør utvikles videre.
Reguleringer og stengte områder
Tekniske reguleringer definerer hvordan, hvor og når fiskere kan fiske. Reguleringene varierer mellom ulike land. Vi anbefaler at hele det tilgjengelige spekteret at tekniske reguleringer brukes. Ved valg av fiskbar størrelse bør denne være koblet til artenes livshistorie. Med data om livshistorie og fangstbarhet kan vi etablere bestandsmodeller som tar hensyn til varierende nivåer av fiskerelatert dødelighet og selektivitet i fisket. Dette kan bli et viktig verktøy når det skal gis råd for å optimalisere innsatsen i fisket og størrelsesreguleringer, samt forutsi mulige endringer i bestandsstørrelse i ulike fiske- og forvaltningsscenarier. Fangstredskapene må utformes slik at de reduserer bifangst og fangst av stor berggylt. Små leppefisk må kunne unnslippe uskadd. Bifangst og undermåls leppefisk skal øyeblikkelig og skånsomt settes ut igjen på fangststedet. Bifangst av andre arter utgjør 10 – 20 % av fangsten (i antall) i de norske fiskeriene. Liknende beregning av bifangst bør gjøres også i andre land. Det trengs studier av overlevelse etter utsetting av bifangst som settes ut igjen på ulike måter. Fisket må ikke foregå i gyteperiodene. Start og lukking av fisket kan variere geografisk og mellom ulike år, så det anbefales at det gjøre et ukentlig prøvefiske for å finne gytetidspunkt. Alternativt kan det brukes en konservativ tilnærming til å sette en fast åpningsdato for fisket. Stengte områder er et godt supplement til andre reguleringer, og sikrer at noen områder har den naturlige sammensetningen av arter og bestandsstørrelser. Stengte områder er også verdifulle som kontrollområder i vitenskapelige undersøkelser av effektene av fisket.
Forvaltningsområder
Det er en fare for overfiske, særlig i områder hvor bestandsstrukturen ikke er vurdert når forvaltningsområdet er definert. Det er viktig at forvaltningsområder er basert på fornuftige biologiske data, slik som genetisk struktur i bestandene. Biologiske forhold varierer mellom artene, så det kan være vanskelig å definere generelle kriterier. Nylige studier av leppefisk viser at lokale bestander er mer avhengig av lokal rekruttering enn av fisk som kommer utenfra. Dette betyr at det kan ta lang tid for overfiskede bestander å bygge seg opp igjen ved at det kommer inn fisk fra omliggende områder. Egg- og larvedrift må derfor inkluderes i beregningene når det defineres forvaltningsområder for de ulike artene. Når denne kunnskapen er på plass, må den formidles til forvaltningsmyndighetene slik at den kan brukes i den fortløpende revisjonen av reguleringer.
Langdistansetransport av fisk
Størstedelen av leppefiskene som brukes er fisket lokalt, men rundt 25% er importert fra Sydøst-Norge eller Sverige. En del av rensefisken kan rømme fra merdene og blande seg med lokale bestander. Flyttinger av rensefisk over lange avstander kan påvirke lokale bestander gjennom innførsel av sykdom og irreversible genetiske endringer. Helsestatus hos rensefisken som flyttes mellom ulike områder er ukjent. Gjentatte transporter øker faren for innførsel og spredning av nye sykdommer. Innførsel av arter som ikke finnes i mottaksområdet fra før, i kombinasjon med sykdomsfremkallende organismer, kan føre til smitteoverføring til nye arter og utvikling av nye sykdommer. Genetiske endringer kan forekomme ved flytting av fisk til områder der det er lav naturlig populasjonstetthet. Langdistanseflytting av leppefisk (i randsonene av artenes utbredelse) bør fra et føre-var-perspektiv unngås. Per i dag er det lov å importere leppefisk fra Sverige under gyteperioden, noe som etter all sannsynlighet øker risikoen for genetisk innblanding. Vi anbefaler at det ikke åpnes for fiske før gytingen er over, og at åpningsdatoene bør være like i Norge og Sverige.
Loggføring av fisketransporter
Norske befraktere behøver ikke å loggføre og rapportere opphavet eller mottakslokalitet for villfanget leppefisk i Norge. På bakgrunn av at de klareste genetiske skillene mellom bestander er innenfor Norges grenser (mellom Skagerrak og nord for Jæren) er det viktig å kartlegge hvor leppefisk som er fisket langs Skagerakkysten havner. Når vi ikke kjenner til opphav og mengder av fisk som fraktes til ulike områder blir det vanskelig å beregne risiko og håndtere problemet med rømminger. Det er behov for data om opphav og utsettingslokalitet for all villfanget rensefisk.
Behov for kunnskap om tapet av rensefisk i merdene
Det er et stort svinn av rensefisk, og ofte få fisk tilbake etter en produksjonssyklus av laks. Forholdet mellom dødelighet og rømming er ikke kjent. Dødeligheten er fra oppdretterne rapportert å være 40% gjennom en produksjonssyklus, men de trodde at den faktiske dødeligheten var nærmere 60 %. Dette misforholdet illustrerer at dagens system for registrering og rapportering av dødelighet av rensefisk ikke er godt nok, gir et dårlig datagrunnlag og gjør det vanskelig å identifisere årsakene til den høye dødeligheten og andelene av fisk som hhv dør og rømmer.
Oppdretterne rapporterte til Mattilsynet at de vurderte avlusing, sykdommer og håndtering som hovedårsakene til dødelighet. Det er ingen felles modell på plass for å overvåke eller kontrollere sykdomsutbrudd hos rensefisk, og svinnet blir ofte ikke oppdaget før lenge etter at det faktisk skjedde.
Det bør derfor etableres systemer for en mer nøyaktig registrering av dødelighet og sykdom hos rensefisk i merdene. Dagens rapporteringssystem for biomasse og dødelighet er utdatert, noe som gjør en analyse av dataene vanskelig. Det haster derfor for både fiskeoppdrettere og myndigheter at det utvikles nye systemer for registrering av dødelighet av rensefisk i merdene.
Forutsetninger for en fortsatt bruk av rensefisk
Mattilsynet har signalisert tydelige krav om at bransjen både må dokumentere at rensefisk kan leve gode liv i merdene og at rensefisk bidrar vesentlig i lusebekjempelsen om de fortsatt skal kunne bruke rensefisk i fremtiden. En tilsvarende konklusjon ble presentert i en nylig rapport fra Rådet for dyreetikk. De anbefaler en mer restriktiv tilnærming enn den myndighetene har varslet. Tilsvarende signal er gitt i Et hav av muligheter – regjeringens havbruksstrategi som ble lansert sommeren 2021. Der står det bl.a. «Den høye dødeligheten, samt helse- og velferdsutfordringene man ser i forbindelse med bruken av rensefisk, gjør det naturlig å sette spørsmålstegn ved om bruken av rensefisk vil kunne fortsette. Hvis næringen i årene som kommer ikke kan dokumentere på en tilfredsstillende måte at rensefisken kan leve gode liv i merdene og bidra vesentlig til lakselusbekjempelse, ser regjeringen det som en naturlig konsekvens at bruken av rensefisk må avvikles.»
For å oppfylle de kravene som myndighetene har formidlet er det nødvendig å vise hvordan det er mulig å gi rensefiskartene et godt liv i merdene, forbedre driftsrutiner og -manualer og fremskaffe data om hvor effektive de ulike artene er. Det må dokumenteres i hvilken grad rensefisk bidrar til å redusere antallet lakselus.
Det må klargjøres om den høye dødeligheten som oppdretterne rapporterer er et resultat av faktorer som kan forbedres eller om rensefisken ikke er i stand til å tilpasse seg forholdene i oppdrettsanleggene (ikke er domestisert).
Specification of species
| NORSK | DANSK | SVENSK | ISLANDSK | SUOMEKSI | ENGLISH | LATIN | |
 | Bergnebb | Havkarusse | Stensnultra | Blettungur | Kivihuulikala | Goldsinny wrasse | Ctenolabrus rupestris |
 | Grønngylt | Savgylte | Skärsnultra | Skerjasnapi | Rantahuulikala | Corkwing wrasse | Symphodus melops |
 | Berggylt | Berggylte | Berggylta | Bergsnapi | Viherhuulikala | Ballan wrasse | Labrus bergylta |
 | Grasgylt | Småmundet gylte | Grässnultra | Kambfiskur | Levähuulikala | Rock cook | Centrolabrus exoletus |
 | Rødnebb/ blåstål | Rødnæb/ blåstak | Blågylta (Blåstråle/ Rödnäbba) | Blettasnapi | Sinihuulikala | Cuckoo wrasse | Labrus mixtus |
") | Rognkjeks | Stenbider | Sjurygg (Stenbit/ Kvabbso) | Hrognkelsi | Rasvakala | Lumpfish (Lumpsucker) | Cyclopterus lumpus |

The scale-rayed wrasse, Acantholabrus palloni, is less common than the other wrasse species in Nordic waters. It lives on deeper waters and usually in rocky or steep areas. It is rarely caught, and little is known about this fish.

The rainbow wrasse, Coris julis, is a doorstep species in Nordic waters. This colourful fish is more common in southern areas but is sometimes observed in Scandinavia. The first observation was two specimens caught in Danish waters in 1834.

Baillon’s wrasse, Symphodus bailloni, is occasionally found in UK waters, but probably not used in aquaculture.
1. Introduction
1.1 A brief history of cleaner fish in salmonid aquaculture
Salmon lice (Lepeophtheirus salmonis) cause great economic losses in the aquaculture of Atlantic salmon (Salmo salar) and rainbow trout (Oncorhynchus mykiss) and have a significant impact on wild populations of salmonids, particularly sea trout (Salmo trutta) that are found in areas with aquaculture activity. Sea lice is directly regulating the production of salmonids in Norway, the world’s largest producer. The Norwegian government has implemented the so called “traffic light system” that manages where the salmon aquaculture industry can grow (green), must stay on the same level (yellow), or must reduce the production (red) for a certain period. This means that a production area given a red light will have a reduction of 6% maximum allowable biomass every second year. The traffic light system is based on regular updated estimations of the impact on wild salmonid populations from infestations of salmon lice emanating from salmon sea cage farms.[1]Ministry of Trade, Industry and Fisheries, 2017. Single sea cage farms are also subjected to a strict salmon lice limit where the average adult female lice per fish should not exceed 0.5, and less than 0.2 during the wild salmon migration period in the spring.[2]Ministry of Trade, Industry and Fisheries, 2012.
The sea lice problem has been present since the start of open pen aquaculture of salmonids in the 1970s. Several drugs have been used for treatment of salmon lice infestations[3]Roth et al. 1993, Burka et al. 1997, Burridge et al. 2010. resulting in the development of drug resistance.[4]Treasurer et al. 2000, Tully and McFaden 2000, Fallang et al. 2004, Sevatdal et al. 2005, Jones et al. 2008, Lees et al. 2008, Besnier et al. 2014, Fjørtoft et al. 2020. Since salmon lice have developed a lack of sensitivity or full resistance towards medical treatments, and such treatments may have a negative impact on wild crustaceans,[5]Urbina et al. 2019. the use of medical treatment to control lice are less favoured now than some years ago.[6]Statistics from the Norwegian Institute of Public Health, https://www.fhi.no/hn/legemiddelbruk/fisk/2019-bruk-av-legemidler-i-fiskeoppdrett/. The concern regarding the environmental impact of chemical and pharmaceutical treatments on non-target organisms[7]Urbina et al. 2019. fuelled the search of alternative methods for de-lousing. This includes various rearing system that fully separate the host from the parasite (e.g., tanks on land or closed units in the sea), near fully (e.g., semi-closed rearing units in the sea with some untreated water exchange with outside water) or only partially (e.g., submerged snorkel-sea cages and preventive skirts around standard sea cages).[8]Barret et al. 2020. It also includes a range of new control methods.[9]Overton et al. 2019. Most prominent of these are various systems for mechanical or thermal de-lousing. This involves first crowding the fish, and then pumping the fish into a treatment system where the lice are either flushed off the fish or heat shocked to lose their grip on the fish from a 20–30 second bath in warm water (28–34 °C). The rearing systems that fully prevent lice infestation are costly and, in many ways, demand a revolution of the industry. Mechanical and thermal de-lousing are also relatively costly, but more importantly represent a significant welfare risk to the treated fish.[10]Overton et al. 2019.
Cleaner fish is one of the first methods developed to combat sea lice in salmon farming. The first record of use of cleaner fish in salmonid aquaculture was recently discovered in a Norwegian newspaper published in 1976, where a salmon farmer reported that he was adding small ballan wrasse (Labrus bergylta) in the salmon cages to reduce lice infestations (mentioned in Bollinger 2020). Although the Norwegian Institute of Marine Research (IMR) was aware of this farmer and contacted him with interest in his approach, the first pilot experiments did not commence until the late 1980s.[11]Bjordal 1988, 1990. Results from the first laboratory-scale trials were promising, followed by successful tests in full-scale production and the method was gradually adopted by commercial scale operations.[12]Bjordal 1992, Treasurer, 1994, Deady et al. 1995, Kvenseth, 1996. In the first years following these trials, interest in the use of cleaner fish was moderate and salmon lice were kept under control using anti-parasitic drugs, but fisheries for wrasse were gradually established in Ireland, Scotland and Norway during the 1990s. Fishers operated inshore and caught wrasses using fyke nets and different types of baited pots (traps).[13]Darwall et al. 1992, Sayer et al. 1993, Varian et al. 1996, Gjøsæter 2002. Five species were targeted: ballan wrasse (Labrus bergylta), corkwing wrasse (Symphodus melops), goldsinny wrasse (Ctenolabrus rupestris), rock cook (Centrolabrus exoletus) and cuckoo wrasse (Labrus mixtus). All these species are found in shallow, hard bottom coastal ecosystems in the North-eastern Atlantic. In addition, the scale-rayed wrasse (Acantholabrus palloni) is found in Scandinavia but prefers deeper waters than the other species and is rarely caught in wrasse fisheries. Baillon’s wrasse (Symphodus bailloni) are occasionally found in UK waters, but their use in salmonid aquaculture is unknown.
During the 1990s, fisheries were not regulated and there is limited information available on landings and catch rates across this period, but it is safe to assume that the fishing pressure was low compared to the 2010’s, at least in Norway. At the same time, trials were conducted on breeding wrasse in both the UK and Norway,[14]Stone 1996, Skiftesvik et al. 1996. but to our best knowledge, no commercial production was established in the 1990’s, and only wild caught wrasse species were used. Most wrasse were locally sourced, but even in 1995 about 500,000 wrasse were translocated from Skagerrak in Southern Norway to farms in Trøndelag, Mid-Norway.[15]Sundt and Jørstad 1998. The interest in cleaner fish slowed during the 2000’s and did not increase until 2009, coinciding with decreasing efficiency of pharmaceutical treatments (Figure 1), but also because cleaner fish was a relatively cheap option for the farmers, who do not have to make significant changes to how they manage and rear their salmon.
By 2016, around 60% of the Norwegian salmon farms reported use of cleaner fish, and this proportion increases to 75% when considering only the coastline south of Nordland county.[16]Lusedata.no. During the initial phase of this “second wave” of cleaner fish interest, farmers still mainly relied on wild caught wrasses, but as demand soared, trials on breeding ballan wrasse and lumpfish (Cyclopterus lumpus) were soon initiated and several commercial producers have been established in recent years.[17]Imsland et al. 2014a-c, 2015a-b, Powell et al. 2018b. Lumpfish were found to be easier to breed than wrasses and are well adapted to colder waters. Juveniles are also effective in de-lousing salmon at low temperatures. At water temperatures below 5-7 °C, several European wrasse species enter a state of reduced physiological activity (torpor), rendering them ineffective for de-lousing purposes.[18]Sayer and Davenport 1996. Lumpfish may therefore be used in northern Norway, where water temperature is too low for wrasses, however their cleaning efficiency may be low during periods with high temperatures and when jellyfish and other preferred plankton, representing alternative food sources, are abundant.[19]Eliasen et al. 2018. Due to these differences in temperature affinity, wrasses and lumpfish are considered complementary cleaner fish, and are used under different conditions. In 2019 the use of cleaner fish in Norway exceeded 60 million fish, where 42 million were lumpfish (Table 1; https://www.fiskeridir.no/Yrkesfiske/Tall-og-analyse/Fangst-og-kvoter/Fangst/Fangst-av-leppefisk and https://www.fiskeridir.no/Akvakultur/Tall-og-analyse/Akvakulturstatistikk-tidsserier/Rensefisk). In comparison, it has been estimated that farmers in Scotland used 480,000 wrasse in 2017 (https://www.sift.scot/wp-content/uploads/2019/02/SIFT-Wrasse-MSP-Briefing.pdf ).
The high demand for cleaner fish in Norwegian farms led to establishment of wrasse fisheries in Sweden in 2010. In 2018, more than 800,000 wrasses were caught at the Swedish west coast and sold to Norwegian farms, predominantly located in Mid Norway.[20]VKM 2019. There are notable differences between Norway and Sweden in terms of fishing practice, management, and scientific survey methodology. The demand for Swedish caught wrasse coincided with the closure of the Swedish west coast eel fishery in 2011, where a large fleet of coastal eel fishermen were left with gear that could catch wrasse. This called for a rapid limitation of the fishing effort in terms of gear and the number of participants in the fishery. Effort limitation and collection of fishery-dependent data has since been at the core of the Swedish management, but a lack of funding has limited the collection of fishery-independent data. Although there has been active collaboration between Swedish and Norwegian wrasse researchers in recent years, especially on mapping the genetic structure (e.g. Faust et al. 2018, Jansson et al. 2017, 2020; Seljestad et al. 2020), there is still a considerable potential for improvement through exchange of knowledge and experiences between scientist and managers across the border, as well as with those in other countries with developing or emerging wrasse fisheries; England, Scotland and Denmark.
Footnotes
- ^ Ministry of Trade, Industry and Fisheries, 2017.
- ^ Ministry of Trade, Industry and Fisheries, 2012.
- ^ Roth et al. 1993, Burka et al. 1997, Burridge et al. 2010.
- ^ Treasurer et al. 2000, Tully and McFaden 2000, Fallang et al. 2004, Sevatdal et al. 2005, Jones et al. 2008, Lees et al. 2008, Besnier et al. 2014, Fjørtoft et al. 2020.
- ^ Urbina et al. 2019.
- ^ Statistics from the Norwegian Institute of Public Health, https://www.fhi.no/hn/legemiddelbruk/fisk/2019-bruk-av-legemidler-i-fiskeoppdrett/.
- ^ Urbina et al. 2019.
- ^ Barret et al. 2020.
- ^ Overton et al. 2019.
- ^ Overton et al. 2019.
- ^ Bjordal 1988, 1990.
- ^ Bjordal 1992, Treasurer, 1994, Deady et al. 1995, Kvenseth, 1996.
- ^ Darwall et al. 1992, Sayer et al. 1993, Varian et al. 1996, Gjøsæter 2002.
- ^ Stone 1996, Skiftesvik et al. 1996.
- ^ Sundt and Jørstad 1998.
- ^ Lusedata.no.
- ^ Imsland et al. 2014a-c, 2015a-b, Powell et al. 2018b.
- ^ Sayer and Davenport 1996.
- ^ Eliasen et al. 2018.
- ^ VKM 2019.
| 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | |
| Ballan wrasse | 270 | 95 | 379 | 1348 | 524 | 577 | 1869 | 681 | 1301 |
| Lumpfish | 431 | 1954 | 3457 | 13385 | 15858 | 30286 | 28986 | 38336 | 36533 |
| TOTAL | 701 | 2049 | 3836 | 14733 | 16382 | 30862 | 30855 | 39017 | 37834 |
Table 1. Sale of farmed cleaner fish to producers of Atlantic salmon and rainbow trout. Numbers in 1000 individuals.

Figure 1: Reported commercial catches of wild wrasses (all species) in Norway in numbers and first-hand value for the fisher during 1998-2020. The reported numbers before 2013 are uncertain since some of the wrasses were reported in weight that were converted to numbers using not-documented conversion factors.
1.2 Life history and reproduction of cleaner fish
Ballan wrasse
Ballan wrasse, the largest wrasse species is a protogynous hermaphrodite, maturing first as female around 22 cm, changing sex when reaching 34–40 cm.[1]Dipper et al. 1977, Darwall et al. 1992, Muncaster et al. 2013. Ballan is the largest wrasse in Nordic waters, with a maximum reported age of 29 years in the UK.[2]Dipper et al. 1977. The males are territorial during the spawning period and provide parental care. There is currently very limited knowledge on how size, age, maturation and sex changes vary spatially for ballan wrasse in Scandinavian waters. Long lived, protogynous hermaphrodites are very sensitive to size selective fishing mortality.[3]Alonzo and Mangel 2004. Given that ballan is the most sought-after wrasse as cleaner fish, there is a clear need for more scientific work on characterizing life history traits in Scandinavian populations. This is especially the case in contrasting areas with different fishing intensity to aid understanding of whether different levels of fishing mortality result in differences in life history characteristics, or if these differences reflect natural variability.
Corkwing wrasse
Corkwing has the shortest life span of the five commercial wrasses, with a maximum age of 3–4 years in South-eastern Norway and South-western Sweden, but they may become considerably older in Western Norway, reaching up to 8 years.[1]Uglem et al. 2000, Halvorsen et al. 2016a. In Norway, corkwing wrasse rarely reach more than 22 cm total length.[2]Halvorsen et al. 2016a. Corkwing wrasse do not change sex, but have clear sexual size dimorphism, which is more pronounced in the western populations. Nesting males grow faster and attain a larger size than females and tend to mature a year later. These males are colourful and defend territories during the spawning season, where they build elaborate nests using coralline algae and other seaweeds. A variable number of males develop as sneaker males, which mimic female morphology, colouration, and behaviour.[3]Uglem et al. 2000. These male morphs are fixed for life – and determined genetically or conditionally during their first year. The sneaker males grow considerably slower than nesting males but mature 1–2 years earlier.[4]Halvorsen et al. 2016a. There are indications that there are more sneaker males in high density populations, where in some cases, they may outnumber nesting males.[5]Halvorsen et al. 2016a.
Goldsinny wrasse
The goldsinny wrasse can attain 20 years[1]Darwall et al. 1992, Sayer et al. 1995a. and is the smallest of the wrasses; it may reach 21.5 cm,[2]T. Larsen, pers.obs. but it is rare to find individuals larger than 16 cm.[3]Halvorsen et al. 2017. Goldsinny males grow faster than females,[4]Olsen et al. 2018. but sexual dimorphism is less pronounced than for corkwing, and seem to diminish in larger age classes.[5]Halvorsen 2017. The growth rate of goldsinny shows high spatial variability on very small scales, which may be attributed to their high site fidelity, where local environmental factors may affect growth rates, such as population density, predation or size-selective fishing.[6]Olsen et al. 2018. The goldsinny males defend territories during the spawning period and are broadcast spawners. Most eggs are pelagic, but it has been observed that a smaller fraction off the eggs sink, which could contribute to local self-recruitment.[7]Hilldén 1984.
Rock cook
There is little published knowledge on the rock cook life history, but a study conducted in Scotland reports a maximum size of 16.5 mm and a maximum age of 8 years.[1]Sayer et al. 1996b. Rock cook does not change sex and males grow faster and live longer than females.[2]Sayer et al. 1996b.
Lumpfish
Despite the high commercial interest, lumpfish are a poorly studied species, and many aspects of their lifecycle and ecology are unknown. Lumpfish are semi-pelagic: the adults are distributed in the open ocean but migrate towards coastal areas to spawn.[1]Holst 1993, Eriksen et al. 2014. This happens during the spring season and the males generally arrive at the coast before the females.[2]Davenport 1985, Kennedy et al. 2016. After an extended courtship, eggs are fertilized by the male as they are laid by the females.[3]Davenport 1985, Goulet et al. 1986. On contact with water, the eggs become sticky and form a clump into a nest. Parental care, which consists in aeration of the eggs and guarding against predators, is undertaken by the male. After laying their eggs, the females seem to rapidly leave the spawning ground.[4]Davenport 1985, Goulet et al. 1986, Mitamura et al. 2012. Females may release two egg batches and spawning occurs over a 4-month period.[5]Gregory and Daborn 1982, Davenport and Lønning 1980, Kennedy 2018. Once fertilized, the eggs hatch 25 to 40 days later depending on water temperature.[6]Collins 1978; Davenport, 1983, Goulet et al. 1988. Newly hatched larvae remain in shallow waters and tide pools until they migrate offshore. It is unknown when or what triggers juvenile fish to leave their coastal habitat. Overall, migration patterns outside of the spawning season are unclear. The age at maturation and longevity for most regions is also currently unknown. Initial estimates for Greenland put age at maturity at 3-4 years old, with the oldest fish examined being 5 years old, which indicates that this species may exhibit a semelparous life strategy.[7]Hedeholm et al. 2014.
Footnotes
- ^ Holst 1993, Eriksen et al. 2014.
- ^ Davenport 1985, Kennedy et al. 2016.
- ^ Davenport 1985, Goulet et al. 1986.
- ^ Davenport 1985, Goulet et al. 1986, Mitamura et al. 2012.
- ^ Gregory and Daborn 1982, Davenport and Lønning 1980, Kennedy 2018.
- ^ Collins 1978; Davenport, 1983, Goulet et al. 1988.
- ^ Hedeholm et al. 2014.
1.3 Geographical distributions
In Norway, all wrasse species are common north to Trøndelag. From there, the abundance of corkwing, cuckoo and rock cook decreases with latitude. An extensive trap survey in Flatanger, northern part of Trøndelag in the late 1990s found only goldsinny and occasional ballan wrasse and cuckoo wrasse,[1]Maroni and Andersen 1996. but corkwing wrasse has recently colonised this area.[2]Faust et al. 2018. Consequently, wrasse used as cleaner fish in farms in mid- to northern-Norway are being introduced from southern populations. Wrasse can escape from the salmon pens through tears in the net, small fish slipping through the mesh.[3]Svåsand et al. 2017, Woll et al. 2013. Recent genetic studies show that cleaner fish in Trøndelag are escaping and reproducing with local populations.[4]Jansson et al. 2017, Faust et al. 2018. Figure 2 shows the northern limits for each species reported in the IMR coastal fyke net survey.[5]K. Nedreaas, unpublished data. These reports have also been checked against species observations done by citizens and reported to the Norwegian Biodiversity Information Centre.[6]www.biodiversity.no.
All five species of wrasse can be found along the Swedish west coast, but in decreasing numbers towards the south, with only the occasional sighting of corkwing and goldsinny wrasse on the Swedish south coast. Only corkwing, goldsinny and Ballan wrasse are caught commercially in Sweden. To this date cleaner fish have not been used commercially in Swedish aquaculture and all cleaner fish are exported to Norwegian farms.
In Danish waters, the only common wrasse species are the goldsinny wrasse and corkwing wrasse – which have highest abundance along the coast of Kattegat. Numbers are slowly declining through the Belts and the Sound and in the Baltic they are rare. All other species are too rare for commercial use, even though they can sometimes be numerous at reefs in the northern Kattegat. Most catches of the other wrasse species are, however, from waters too deep for commercial wrasse fishing.
In UK waters all five species of wrasse are common and are caught commercially. Baillon’s wrasse Symphodus bailloni is also found on the south coast of the UK, but its use in commercial Scottish aquaculture is unknown. In the UK, commercial fishing occurs in Scotland and on the south coast of England. Wrasse from England are transported to Scotland for use in Scottish aquaculture.
Lumpfish are mostly present in the Nordic Seas, along Newfoundland, Greenland, Iceland, Norway, and Russia. They are also found in areas of lower salinities such as the Baltic Sea and Hudson Bay.[7]Davenport 1985. Southern limits of their distribution area are Chesapeake Bay on the western side and the English Channel on the eastern side.
1.4 Genetic structure
Ballan wrasse
Knowledge of the genetic structure of Ballan wrasse populations has recently been updated. In a study by Seljestad et al. (2020), more than 1000 fish from 19 locations were genotyped for 82 SNPs. A clear genetic break dividing Scandinavian populations into north western (NW) and south-eastern (SE) groups, was identified. The division coincides with the one previously identified for corkwing wrasse,[1]Blanco González et al. 2016. and is located on the coast of southern Rogaland, in southwestern Norway. This area constitutes the longest continuous stretch of sandy bottom substrate along the coast (the Jæren beaches ~26 km), and likely also a natural barrier for gene flow for ballan wrasse, which needs hard bottom substrates for their sticky benthic eggs. Previous studies have concentrated on population divergence, history and amount of genetic variation on a larger scale based on mitochondrial sequence data,[2]Almada et al. 2016; D’Arcy et al. 2013. and included samples from southwestern Norway, around the British Isles, and along the coastline of France, Spain and Portugal as well as from the Canary Islands and Azores. Both studies revealed a significant decrease in genetic diversity towards the north and existence of clear genetic structure along the species distribution area. Lower genetic variation in Norway compared to Spain was also observed in the study by Seljestad et al. (2020), and in another study Casas et al. (2021). The lower variation in Scandinavia is likely due to one or multiple founder events when populations have gone through bottlenecks during the (re)colonization from southern latitudes after the last glacial period. Seljestad and colleagues (2020) suggest that the main route of colonization of Ballan wrasse into the Scandinavia could have been via the British Isles and across the North Sea to the Norwegian west coast and then further on towards north and south. This is supported by the observation that the genetic differentiation between SE Scandinavia and Spain was lower than between NW Scandinavia and Spain. Studies on the two main morphs, spotted and plain, have reported significant differences in growth, mortality, and investment in reproduction.[3]Villegas-Ríos et al. 2013a and b. As these morphs occur in sympatry, it is possible that there are genetic differences between the morphs. A study using microsatellite markers on Spanish populations[4]Quintela et al. 2016. discovered a large enough genetic difference between these two morphs to suggest that they could represent cryptic species. However, another study on three mitochondrial and one nuclear genetic marker did not find any significant difference.[5]Almada et al. 2016. Seljestad et al. (2020) studied this matter further and expanded upon the Quintela et al. (2016) work by analyzing phenotyped samples from multiple locations with a panel of SNPs. Besides the same samples from Galicia, Spain as used in the Quintela et al. (2016) study, they included both types of fish (and intermediates) from four locations in Scandinavia (Smøla, Flødevigen, Strömstad and Gothenburg). They confirmed clear genetic differentiation between the phenotypes in Spain suggesting that in the south, spotty and plain colour morphs represent distinct genetic groups. On the contrary, no genetic differentiation was detected between the sympatric phenotypes in any of the four Scandinavian samples, and the phenotypic variation was observed to be less distinct. These results are in line with the recent study by Casas et al. (2021) in which fish of both morphotypes collected in Spain, France, Ireland, and Norway were characterized with 39 602 genome-wide SNPs. The authors found strong support for the genetic differentiation of plain and spotted individuals in Spain, moderate support for France and Ireland, and a complete mix of both phenotypes in Norway. The biological role of these morphs, and the underlying mechanisms upholding them, remain unclear, however. The ballan wrasse genome was sequenced by Lie et al. (2018) and is available at the European Nucleotide Archive (accession number: PRJEB13687).
Corkwing wrasse
Several studies of corkwing wrasse have discovered a significant reduction in genetic diversity in northern Europe, as shown for ballan wrasse.[6]Robalo et al. 2012, Knutsen et al. 2013. This reduction of genetic diversity has been coupled with a large genetic break between Scandinavian and UK populations. The genetic break and reduced genetic diversity is believed to be a historic effect of the species going through one or multiple bottlenecks when colonising the Scandinavian coast.[7]Robalo et al. 2012, Knutsen et al. 2013, Mattingsdal et al. 2020. The lack of suitable rocky habitats between Scandinavia and the Atlantic in combination with a short pelagic phase would also explain the lack of gene flow and how the genetic break has been maintained since colonisation. There is also a strong population structure within Scandinavia. Blanco Gonzalez et al. (2016) discovered a second major genetic break separating populations in Skagerrak from populations along the west coast of Norway. They hypothesised that a stretch of non-suitable habitats of sandy beaches are acting as an environmental barrier to gene flow, preserving genetic structure on a small geographical scale. The identified break aligns with considerable differences in life history traits, where Skagerrak corkwing populations have been found to grow faster and mature earlier than the populations further north[8]Halvorsen et al. 2016. and can thus have important implications for management. Genomic analysis of the demographic history of corkwing wrasse suggest that Scandinavia was colonised from the British Isles around 11 thousand years ago, around the time of de-glaciation.[9]Mattingdal et al. 2020. The Skagerrak population was either: 1.) founded from the western Norway population around 10 thousand years ago and has largely been isolated since the split; or 2.) was not colonised from western Norway but from a different glacial refuge and current gene flow across the break is a result of secondary contact. North Sea populations might contain further regional substructure. A study using microsatellite genetic markers discovered a pattern of isolation by distance along the Norwegian west coast.[10]Blanco González et al. 2016. The presence of small fjords was suggested to be responsible for the regional pattern of genetic structure. New data suggest another genetic break along the western Norway coast, although much less distinct.[11]Faust et al. 2021. Within Skagerrak, genetic population structure appears less clear. Knutsen et al. (2013) found there to be “a tendency for substructure”, whilst Blanco Gonzalez et al. (2016) maintained that Skagerrak populations are genetically homogeneous. An ongoing genomic study by Faust and colleagues indicates that there is little to no structure within Skagerrak and Kattegat, suggesting either high long-distance gene flow, a severe population bottleneck and/or a very recent population origin. An earlier study by Faust et al. (2018) discovered that roughly one third of wild corkwing wrasse caught in Trøndelag originated from or were the first- or second-generation offspring from corkwing caught in Skagerrak. This suggests that corkwing wrasse not only escape from fish farms but will also hybridize with local populations, if present. However, the study only investigated a small number of fishes caught in one area. Further investigation to the geographical extent and magnitude of escapees indicates that hybridization and introgression from Skagerrak is mainly noticeable at the northern limit of the distribution range where local populations are smaller.[12]Faust et al. 2021. The question remains if the lack of escapees and hybrids in more southern locations is due to less escapees, lower survival or to sampling effort by population size. It is also possible that escapees do better in smaller populations due to less competition for food, nesting grounds and mates. Given the known differences in growth and maturation between the North Sea and Skagerrak populations, we can expect to see changes not only in genotype composition but also phenotype and potentially fitness of local populations. The corkwing wrasse genome was sequenced by Mattingsdal et al. (2018) and is available at the NCBI Sequence Read Archive as part of Bioproject PRJNA354496.
Goldsinny wrasse
Like other wrasses, the goldsinny wrasse populations are clearly genetically divided across the North Sea[13]Jansson et al. 2017; 2020. and shows a trend of reduced genetic variation towards the northern areas. In contrast to the corkwing and ballan wrasse, which both show highly differentiated populations in Scandinavia with a strong genetic break, goldsinny has a relatively weak population structure, but with a clear pattern of isolation-by-distance.[14]Jansson et al. 2017. The difference in population structure between the species could be related to differences in population connectivity caused by distinct reproductive strategies. In contrast to the benthic eggs in the other cleaner fish, goldsinny is the only Nordic species releasing planktonic eggs.[15]Darwall et al. 1992, Potts 1985, Hillden 1984. However, similarly to corkwing wrasse, goldsinny populations in farming areas in Trøndelag display a relatively low genetic differentiation from wild populations in Skagerrak, indicating that they may also escape and mix with local populations.[16]Jansson et al. 2017. The goldsinny wrasse genome was sequenced by Jansson et al. (2020) and raw reads are available at the NCBI Sequence Read Archive (Bioproject PRJNA508986).
Footnotes
- ^ Blanco González et al. 2016.
- ^ Almada et al. 2016; D’Arcy et al. 2013.
- ^ Villegas-Ríos et al. 2013a and b.
- ^ Quintela et al. 2016.
- ^ Almada et al. 2016.
- ^ Robalo et al. 2012, Knutsen et al. 2013.
- ^ Robalo et al. 2012, Knutsen et al. 2013, Mattingsdal et al. 2020.
- ^ Halvorsen et al. 2016.
- ^ Mattingdal et al. 2020.
- ^ Blanco González et al. 2016.
- ^ Faust et al. 2021.
- ^ Faust et al. 2021.
- ^ Jansson et al. 2017; 2020.
- ^ Jansson et al. 2017.
- ^ Darwall et al. 1992, Potts 1985, Hillden 1984.
- ^ Jansson et al. 2017.

Figure 2. Northern natural geographical limits for (A) Corkwing wrasse (Symphodus melops) and Rock cook (Centrolabrus exoletus) (N 64°50.3’), (B) Ballan wrasse (Labrus bergylta) and Cuckoo wrasse (Labrus mixtus) (N 66°30’), and (C) Goldsinny wrasse (Ctenolabrus rupestris) (N 69°29.4’).
Lumpfish
Across the lumpfish’s distribution range, three distinct genetic groups have been proposed: West Atlantic, Iceland & East Atlantic, and the Baltic Sea.[1]Pampoulie et al. 2014. A finer-scale study along the coast of Greenland found a pattern of isolation by distance, but only within the group as all samples were more closely related to North American samples than Icelandic ones, regardless of geographic distance.[2]Garcia-Mayoral et al. 2016. On the contrary, Jónsdóttir et al. (2017) found no signs of spatial genetic structure among lumpfish studied along the Norwegian west coast. These large homogeneous groups suggest long distance migration within groups and strong barriers between groups. It has been proposed that lumpfish in the West and East Atlantic are separated by cold southward polar currents, and that populations in the Baltic Sea likely became isolated during the Last Glacial Maximum.[3]Pampoulie et al. 2014. These together explain the observed major genetic groups and large-scale patterns. However, a recent study proposed that there might be finer genetic structure with significant differences on a much smaller, regional scale.[4]Whittaker et al. 2018. This is likely to be due to homing behaviour during spawning that leads to decreased migration between populations.[5]Kennedy et al. 2015, 2016. The study by Whittaker et al. (2018) found that Lumpfish in Iceland and the English Channel were significantly differentiated from each other and from all other East Atlantic samples. Furthermore, they found that one of three sample locations in Norway was genetically different from other East Atlantic samples, suggesting that there might be previously undiscovered substructure along the Norwegian west coast. Such discrepancies between studies could be explained by sampling scheme and coverage, as well as by selection of the used genetic markers. In all studies mentioned above, a relatively small set of traditional genetic markers (microsatellites) was utilized. As it has been shown that more markers usually lead to better resolution,[6]Putman and Carbone 2014, Whitaker et al. 2020. the genetic population structure of lumpfish globally and within Scandinavia is currently being revised in an ongoing genome-wide study by Jansson, Faust and colleagues.
2. The ecological impacts of fishing and using wrasse as cleaner fish
This chapter reviews each country’s scientific monitoring programmes of the wrasse stocks for obtaining knowledge on the population dynamics and impact of fishing on wrasses, and which is the foundation for management advice (chapter 3). Further we provide an overview of known and potential impacts of fishing and using wrasse, ranging from any direct impacts from fishing on the wrasse stocks, via the indirect ecosystem effects and lastly the consequences of long-distance translocation and escapement of wild-caught wrasse in distant regions.
2.1 Stock monitoring
The monitoring of abundance, size structure and distribution of exploited stocks is a necessity for sustainable fisheries management. This allows scientists to track changes in the populations and assess the impact of fishing and other environmental factors. Both fishery independent and fishery dependent surveys can be used for this purpose. Fishery independent surveys carried out by scientific personnel are generally more expensive but can provide more accurate and detailed data than possible to obtain from the fisheries. Catch per unit effort (CPUE; Catch/effort) is a relative abundance index, which under some circumstances can be assumed to be proportional to absolute abundance. Thus, changes in CPUE with time could indicate stock status, and therefore serve as a measure to advise adjustment in fishing effort. However, using CPUE as an abundance index has several shortcomings and pitfalls, especially for sedentary coastal fish such as wrasse: First, there is high spatial heterogeneity in abundance, species composition, size structure and life history traits, at both local and regional scales. They have very high site fidelity and deeper waters (> 50 m) are habitat barriers,[1]Halvorsen et al. in review. which means that spatial variability in environmental conditions and habitat quality has a strong influence on demography and population dynamics in local populations. Second, the spatial fishing effort is also highly variable within and between areas, so it is not possible to draw inferences on the impact of fishing on stocks without knowledge of the past fishing pressure in each location. Third, catchability of wrasses is highly dependent on temperature, season, depth, exposure,[2]Halvorsen et al. 2020a, b. meaning that catch data without such environmental covariates are likely to yield biased abundance indicators.[3]Maunder and Punt 2004. The activity of the fish is also closely related to the temperature in the sea and more generally to the season. Fishery independent surveys are also prone to these biases if not accounted for. To assess the impact of fishing, surveys conducted simultaneously in protected and exploited sites is a well-established approach for species with limited mobility, ideally to be done before and after fishing or protection has been conducted (BACI-design, Before-After-Control-Impact).
Solutions adopted in Norway
Fishery dependent surveys: In Norway, IMR has gathered catch data from a selection of reference fishers from 2011-2020. This has been used to calculate CPUE in different management regions, but due to limitations in the surveys design in 2011-2018, it was not possible to standardize the data and therefore establish a reliable time series from the data in this period. This is partly because the fisheries have undergone major changes in fishing gear and the sizes kept which means that both the sampling gear and the sizes included in the catch estimators have not remained the same. Until 2016, fishers were instructed to report the number of retained wrasse and the numbers of gears used per day. The number of discarded fish was reported from a subset of the gears (5-10 %), thus the sampling units for retained and discarded fish were not identical. CPUE was calculated as the total catch divided by the numbers of gears, and then standardized to 24 hours soak time. However, this introduced an erroneous assumption of a linear proportional relationship between number fish and soak time – but trap saturation seems to happen early in the capture process and change little after the first hours.[4]Halvorsen et al. 2020a, b. Furthermore, length was not measured in this survey, only the minimum retained size, and it was therefore not possible to know the size component that makes up the retained and discarded CPUE index. Fishers are known to change the criterion for retainment throughout the season. From 2017, both retained and discarded fish were counted in the subsampled gear, allowing for estimating CPUE combined for discard and retained fish from this subsample.
An internal review of the survey methodology was conducted in December 2018. The challenges summarized above was identified and it was decided that fishers should count and measure length on all wrasses in two standardized pots per day. All by-catch is also identified to species level and counted, and lobster and cod is length measured. Fishers are provided with a custom-made device for measuring wrasses. The pots would be fitted with a Star-Oddi data logger that records depth, temperature, and time. The fishers are also instructed to report an accurate GPS position for each trap. These data can then be included as fixed covariates when analysing CPUE in a generalised linear mixed effects model (GLMM) with trap as the sampling unit, and fisher and day as random effects. The preliminary analysis shows that such a model reduces inter-annual and inter-fishery variance in the CPUE estimates to a much larger degree than when CPUE is measured on an aggregated sample from a single day.[5]Halvorsen et al. 2020a, b.
IMR has not been running a national scale scientific survey to monitor wrasse populations in Norway. For regional trends, the fishery-dependent survey has been assumed to be a more cost-efficient and logistically feasible methodology to obtain the necessary spatial and temporal coverage in the data. However, several location-specific surveys are being (and have been) carried out which provide more in-depth knowledge on population dynamics on restricted spatial scales. For example, abundance, size, and age was compared with data from a fyke net survey conducted in and outside MPAs in Southern Norway in 2013,[6]Halvorsen et al. 2017. while a tagging study in 2014 was successfully used to assess fishing mortality on corkwing wrasse in Western Norway.[7]Halvorsen et al. 2016b. IMR has also established more long-term time series. In the period 2013-2020, IMR conducted annual fyke net and trap surveys in specific sites in Austevoll and Flødevigen. Until 2018, weekly sampling was conducted for 6-9 weeks in May-July, primarily to monitor the duration of the spawning period and gather knowledge on temporal fluctuations in abundance and size structure. For several years, the results were reported to the Directorate of Fisheries on a weekly basis, which opened the fishery when the spawning season was nearly finished. The survey was not continued when the management settled on a fixed opening date from 2018, but a new seasonal fyke net survey with some spatial overlap was established in Austevoll from 2017 in and outside marine protected areas where commercial fishing is prohibited. In 2019 and 2020, an intensive experimental fishery was conducted in one of the islands inside the MPAs where the fyke net survey is conducted in Austevoll. This was done to assess how populations are impacted by a fishery of known intensity. All commercial wrasses are PIT tagged, allowing for estimating natural mortality, fishing mortality, catchability, gear selectivity and movement.[8]Halvorsen et al. in review.
Solutions adopted in Sweden
Landings per unit effort (LPUE) is currently the main variable for estimating stock development of the wrasse species. The Swedish Agency for Marine and Water Management (SwAM) collected landings data from fisher’s logbooks between 2010 and 2013. Since 2014 it is mandatory for all fishermen to keep an extended record of which days fishing takes place, the number of times gear has been emptied in a day, areas fished, and the amount and type of gear used. For the target species, the reporting is limited to the sizes and specimen landed and may be influenced by changes in the demand for cleaner fish. This fishery dependent data covers the entire Swedish fleet, but otherwise suffers from very similar limitations as described for the earlier Norwegian data.
For the information to become more useful in relation to spatial management, it would be desirable to use accurate, fine-scale fishing positions. In some cases, only the broad-scale area of fishing is reported, which makes it difficult to analyse the spatial distribution of the fishery. Fishermen rarely separate landings by gear in their logbooks when different gears are used simultaneously. This is problematic as the efficiency of the gear types (LPUE) differs for all three species.[9]Halvorsen et al. 2017. Corkwing and ballan wrasse are mainly caught in fyke nets, whilst pots are more efficient in catching goldsinny wrasse. There is also a difference in when the different species are being targeted; ballan wrasse is mainly targeted in May and June, whilst corkwing and goldsinny are mostly targeted from July until October. Therefore, a separate account of landings from pots and fyke nets would be preferred. It is also important that the fishery indicates changes in the design of the gear on an annual basis, as well as changes in the choice of target species by fishing trip.
In the extended record, the number of individuals of the three target species retained and bycatches of cod and eel are reported. For the target species, this means that only individuals within a certain size fraction of the catch are reported in the logbook entry. However, no information on undersized fish, spawning status and/or injured fish is obtained as these are released. In addition, there is a need for information on additional bycatch species other than cod and eel. Furthermore, in the logbooks landings is reported by weight.
In Sweden, The Swedish University of Agricultural Sciences (SLU Aqua) carries out annual fyke net surveys in August and October in reference areas to monitor long-term trends of fish communities. There are two such reference areas along the Swedish west coast archipelago where wrasses are monitored: Fjällbacka with annual sampling since 1989 in October, and since 1998 in August and Älgöfjorden in Marstrand with sampling since 2002. The existing analysis of the spatial distribution of the wrasse fishery indicates that little to no fishing has occurred in the reference areas. The two survey locations have great value as reference areas for changes in stocks caused by large scale human influence and natural variability. However, it is less clear whether the surveys can reflect stock changes caused by fisheries because of the limited movements in these species.[10]Halvorsen et al. 2021. Furthermore, the timing of the survey and the locations are unfortunate for ballan wrasse stocks as this species has rarely encountered even before the fishery started. The fyke net data are therefore presently not used for the management of the wrasse fishery.
To study if Swedish wrasse populations show signs of depletion, such as lower density or average size in fished areas, a survey was conducted comparing a 2.6 km² large marine protected area with two equally large, fished areas. A total of 112 pots and 164 fyke nets were deployed over a nine-day period in August 2016. Data showed no clear evidence of depletion for any of the three species of wrasse targeted in Sweden, ballan, corkwing and goldsinny wrasse.[11]Bourlat et al. 2021. However, it should be noted that overall, very few ballan wrasse were caught in both areas making it difficult to assess the effect of fishing on this species. To get a better overview of the stock, and get a representative length distribution over available fish, pilots for a new survey tailored to monitor ballan wrasse are currently being tested.
Solutions adopted in Denmark
No data from fishery-dependent surveys are available from Denmark since no commercial fishing for wrasses has been done. Since 2019, however, a collaborative EHFF project including fishers, a consultancy company and the University of Copenhagen has collected fisheries data from explorative fishing. The project monitors the CPUE in both traditional wrasse traps and seal-resistant traps.
Knowledge about wrasses in Danish waters is very limited since the shallow waters along the coasts have not been regularly monitored. The current knowledge from The National Danish Fish Atlas was recently published online.[12]Carl and Møller 2019. Most information has been obtained by e.g. eel traps[13]Støttrup et al. 2018. and snorkel surveys.[14]Sigsgaard et al. 2017; Holm-Hansen et al. 2019. Some information also came from local gill-net surveys e.g. from restoration of reefs[15]Støttrup et al. 2014. and/or camera-based surveys from wind farms.[16]Hansen et al. 2012. Finally, in recent years a few studies have used environmental DNA for detection and abundance estimates.[17]Sigsgaard et al. 2017.
Experimental fishing with traditional commercial wrasse pots has been conducted in four different locations, ranging from high salinity (approximately 32 ‰) waters in the northern part of Jutland (Skagen) to low salinity waters (approximately 12‰) near Copenhagen (Øresund/Kastrup). The fishery was conducted over a three-month period from September to November 2019. In total 9,273 wrasses were caught. DNA samples were also collected to determine if there is genetic variation between wrasses from the four locations. This is a step towards getting an export permit for wrasses to Norway. The two main species caught in the experimental fishery in 2019 were goldsinny wrasse and corkwing wrasse, which make up 99.9% of the total catch, with the distribution of 47% and 53% respectively. Only 10 individuals of ballan wrasse were captured in the four locations during the three-month period. The results from the experimental fishery indicate a distinct difference in size distribution between the four locations. In Skagen 92% of the wrasses were above 11 cm, while only 34% in Kastrup. The variation may be due to the difference in salinity, as the inner Danish waters are not likely to be an optimal habitat for wrasses. It is unknown whether they are just short lived, or if they migrate to areas with higher salinity as they get older.
The National Danish Fish Atlas has assessed the occurrence and abundance of wrasses from snorkel surveys at numerous locations along the Danish coastline. The results indicate that wrasses are present in large numbers along almost every pier and around rock formations. In 2020 and 2021 the focus will be on gathering information regarding by-catch, spawning periods and fish behaviour in and around the pots, to determine the catch efficiency of the commercial wrasse pots.
Solutions adopted in the UK
In southern England, the Inshore Fisheries and Conservation Authorities (IFCAs; the management authorities for inshore fisheries in England) take varied approaches to monitoring stocks of wrasse. In the Cornwall and Devon & Severn IFCA districts annual fishery-dependent surveys of the live wrasse fishery are conducted to generate evidence to support adaptive management. These surveys began in 2016 in Cornwall and 2017 in Devon & Severn. During these on-board observer surveys of commercial fishing operations, IFCA officers record the date, time, bait type, fishing gear soak time and fishing locations (start and end points of each string of pots). Each wrasse caught is identified to species level and total length measured to the nearest 0.5 cm. The granularity of the data varies slightly between IFCA districts. Cornwall IFCA record the catch on a ‘trap by trap’ basis, recording each trap as a separate sampling unit, whereas Devon and Severn IFCA, pool the recorded metrics of fish caught from all traps along the string. From these surveys, both total catch per unit effort (CPUE; all fish caught regardless of whether retained or returned) and landings per unit effort (LPUE; only fish that are retained) can be calculated. Southern IFCA have completed a limited number of opportunistic fishery-dependent on-board surveys but rely mainly on the submission of catch return forms from the fishers to monitor fishing effort.
Each IFCA district requires fishers to submit fishing returns forms, where fishers record the number of wrasse retained, the type and amount of fishing gear used, and broad-scale fishing location for each day fished. In the Southern district, this is currently done by the fishers on a voluntary basis, as was the case in Cornwall and Devon & Severn before the introduction of byelaws that included catch return forms as a permit condition in 2019 and 2017, respectively. From this the IFCAs can calculate LPUE, however this measure is not as detailed as the LPUE calculated from the on-board observer data.
These surveys and data collection have been used not only to expand the knowledge base of wrasse ecology and distribution in South-West England,[18]Henly et al. 2021; CIFCA 2017 report. but also to gather an evidence base to inform the adaptive management of the fisheries. For example, using data from fishery-dependent observer surveys, Curtin et al. (2020) highlighted significant declines in both CPUE and LPUE of rock cook between 2017–2019 in the Devon & Severn IFCA’s (DSIFCA) district. On this basis, DSIFCA altered the potting permit byelaw to prohibit removal of rock cook from the fishery.[19]DSIFCA, 2020.
Footnotes
- ^ Halvorsen et al. in review.
- ^ Halvorsen et al. 2020a, b.
- ^ Maunder and Punt 2004.
- ^ Halvorsen et al. 2020a, b.
- ^ Halvorsen et al. 2020a, b.
- ^ Halvorsen et al. 2017.
- ^ Halvorsen et al. 2016b.
- ^ Halvorsen et al. in review.
- ^ Halvorsen et al. 2017.
- ^ Halvorsen et al. 2021.
- ^ Bourlat et al. 2021.
- ^ Carl and Møller 2019.
- ^ Støttrup et al. 2018.
- ^ Sigsgaard et al. 2017; Holm-Hansen et al. 2019.
- ^ Støttrup et al. 2014.
- ^ Hansen et al. 2012.
- ^ Sigsgaard et al. 2017.
- ^ Henly et al. 2021; CIFCA 2017 report.
- ^ DSIFCA, 2020.
2.2 The effects of fishing on wrasse populations
The various monitoring efforts reviewed above and other dedicated studies has provided important insights in how the fisheries affects the wrasse populations. However, many knowledge gaps still remain, especially since both management regulations and fishing practice has changed considerably since the fishery started and varies between countries and regions.
The wrasse fishery has differential impact on the various target species, influenced by each species unique life history traits, habitat preferences and catchability in the gear. Corkwing and ballan are the species with the highest overlap with the fishery (0-6 m), while the density of goldsinny appears to be constant from 0 to 20 m depth.[1]Halvorsen et al. 2020a; Figure 3. This implies that corkwing and ballan have the highest probability of encountering fishing traps.
Case studies have shown that fishing can affect local populations. A tagging experiment with corkwing wrasse in Austevoll, western Norway revealed that fishing mortality was considerable; up to 40 % of the tagged individuals were harvested over a two-month period.[2]Halvorsen et al. 2016b. Nesting males had higher capture probability than females, so the fishery may be sex selective, which can have negative effect on parental care and egg survival.[3]Kindsvater et al. 2020. On the Skagerrak coast, marine reserves had higher catch per unit effort of goldsinny and corkwing wrasse compared to nearby areas open for fishing.[4]Halvorsen et al. 2017. In contrast, a recent master thesis analysed a decade with scientific survey data in and outside a Marine reserve on the Skagerrak coast and found no clear differences between protected and fished areas.[5]Reamon 2020. However, as in this and the other studies, the fishing effort was not measured in the open areas. A few UK studies reports declines in CPUE and effects of size structure of goldsinny,[6]Sayer et al. 1996a; Varian et al. 1996. corkwing[7]Darwall et al. 1992. rock cook[8]Curtin et al. 2020. and ballan[9]Henly et al. 2021. following the establishment of locally intensive fisheries on the British Isles. The Swedish fishery is local and small-scale; fishermen can only fish for wrasse after receiving an exemption for using gear, such as fyke nets and pots, without escape openings for lobster. Roughly 14 such exemptions have been handed out for wrasse fishery annually. In 2016 SLU Aqua and Gothenburg university conducted a survey comparing fished and unfished areas on the Swedish west coast. There were no clear differences between the number or size of the fish in the two areas.[10]Bourlat et al. 2021. It should be noted that only a few ballan wrasse were caught overall and thus this species could not be properly evaluated. Currently, Sweden has no management plans that connect exploitation rates with the biology of the different wrasse species.
The impact on the various species depends both on the technical regulations and the behaviour of the fishers. Norwegian fishers report that the majority of ballan, corkwing and goldsinny above the minimum size limit are retained (Figure 4). The minimum size limit for ballan (140 mm) has little effect, hence 97 % of the catch of this species is reported to be retained by fishers, while for corkwing and goldsinny, ~ 70 % are retained. The demand for rock cook and cuckoo wrasse as cleaner fish is very low, and the majority are released. In Norway, the fishery is now managed with individual quotas for all fishers, in addition to a maximum catch set for three management regions.[11]Halvorsen et al. 2020b. Although quotas can be effective in reducing the overall fishing intensity, they are not species-specific, which means that the species composition may reflect what species the fishers target and retain to an unknown degree. The price per individual for ballan wrasse is higher and has increased more than the other species in recent years (Figure 5), which should provide an incentive for fishers to increase the share of ballan in their quota. Thus, quotas induce a risk of overfishing for the more vulnerable and/or economically valuable species, in this case the ballan wrasse.[12]Halvorsen et al. 2020b.
Lastly, the vulnerability to fishing is highly dependent on the life history traits and reproduction of species. Knowledge of life history traits help us to understand how populations respond to harvesting and to implement informed management measures to avoid overfishing.[13]Kindsvater et al. 2016; 2020. A general principle is that populations with short life cycles (early maturation, short life span) are more resilient to exploitation than those with late maturation and long lifespan. Wrasses display high variability in life history traits at both regional and local scales but there are still considerable knowledge gaps. For example, information on growth rate, life span, size/age at maturity and sex change is lacking for ballan wrasse in most areas where it is harvested. Furthermore, studies on the importance of nesting males in reproduction (ballan, corkwing) is warranted, as males appears to be disproportionally targeted due to their larger size at age.[14]Halvorsen et al. 2016a. Routine, standardized sampling programmes of scales and/or otoliths should be established in key areas, ideally in conjunction with sampling for genetic structure, as life history traits may differ among genetically isolated populations. Protocols for sampling should be similar across countries (gear type, sampling time, sexing etc), and data should be shared to facilitate for a broader understanding of the underlying mechanisms for life history variability.
Footnotes
- ^ Halvorsen et al. 2020a; Figure 3.
- ^ Halvorsen et al. 2016b.
- ^ Kindsvater et al. 2020.
- ^ Halvorsen et al. 2017.
- ^ Reamon 2020.
- ^ Sayer et al. 1996a; Varian et al. 1996.
- ^ Darwall et al. 1992.
- ^ Curtin et al. 2020.
- ^ Henly et al. 2021.
- ^ Bourlat et al. 2021.
- ^ Halvorsen et al. 2020b.
- ^ Halvorsen et al. 2020b.
- ^ Kindsvater et al. 2016; 2020.
- ^ Halvorsen et al. 2016a.
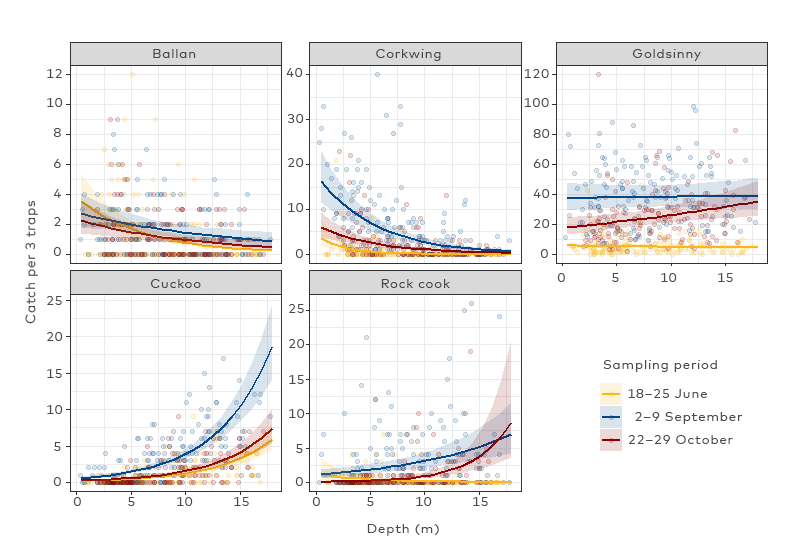
Figure 3. Catch as a function of depth for the five wrasse species in pots in three different time periods. The points are the observed numbers per sampling unit (set of three traps), solid lines are model predictions, standardized for soak time and wave exposure, with confidence interval (95 %). Figure adopted from Halvorsen et al. 2020a)
Figure 4. Length distribution of five wrasse species in the Norwegian fishery. Fill indicate the proportion retained (blue) and released (yellow) in each 1 cm length interval. The vertical dashed line shows the minimum size limits. Data from all reference fishers in 2019-2020 (N = 1535 pot hauls).

Figure 5. Average first-hand price to fisher (in NOK, upper panel) per wrasse species during 2013–2019. Wrasse landings in Norway and Sweden 2013–2020 (million fish, lower panel). Source: The Norwegian Directorate of Fisheries and Swedish Agency for Marine and Water Management. Some catches of cuckoo wrasse are also registered in the same period.
2.3 Wider ecological impacts of wrasse fisheries
If wrasse fisheries change species composition, size structure of nearshore ecosystems, this may in turn affect functional ecology and trophic interactions. Wrasses are predominantly found on hard bottom, macroalgae covered habitats, where they find shelter, food and nesting sites.[1]Costello 1991; Thangstad 1999; Skiftesvik et al. 2014. The wrasses have high site fidelity and limited home range.[2]Sayer 1999; Villegas-Ríos et al. 2013; Halvorsen et al. 2016b. Wrasses are mesopredators, feeding on sessile or slow moving prey, such as crustaceans, molluscs, gastropods, hydrozoans and polychaetes.[3]Alvsvåg 1993; Deady and Fives 1995a,b; Sayer et al. 1995a, 1996b, Bourlat et al. 2021. Wrasses are important prey for larger piscivores,[4]Steven 1933; Rui Beja 1995; Nedreaas et al. 2008. except for the adult ballan and cuckoo wrasse which have fewer natural predators due to their larger size. Due to their high abundance and biomass in shallow water ecosystems, wrasses can be regarded as key species. Despite several studies on diet of wrasse, many of these show large variations in time and space within species, indicating an opportunistic feeding behaviour reflecting local prey availability.[5]Alvsvåg 1993; Deady and Fives 1995b,a; Sayer et al. 1995b.
However, there are few published studies on the ecological links between wrasses and other ecosystem components that go beyond correlations and descriptions. In Southern Scandinavia, the abundance of wrasse has shown increasing trends the last 30 years, which has coincided with the decline of large piscivores, such as the coastal cod.[6]Eriksson et al. 2011. In the same timeframe, the ephemeral algae have substituted perennial macroalgae in many areas. Higher wrasse abundance may have led to increased predation on important grazers on ephemeral algae, and it has been hypothesized that commercial wrasse fisheries could have a positive effect on coastal food webs, with the fishery taking the predatory role of the depleted piscivores.[7]Östman et al. 2016. An experiment that lasted over 15 days found that total invertebrate abundance was decimated by 61% in the presence of wrasse and by 36% in the tanks without wrasse (Bourlat et al. 2021). This also resulted in significantly less fouling by epiphytic bryozoans on algae in the tanks without wrasse, suggesting cascading effects may result if wrasses are decimated by fishing. However, wrasse do not only feed on mesoherbivores, but also on intermediate consumers, and also primary producers, which indicates that wrasses may
have multiple effects on food web dynamics. A recent paper estimated the that the number of wrasses eaten by cormorants in Norwegian waters is comparable to the fisheries catches. This highlight a potential conflict between wrasse fisheries and bird populations.[8]Dehnhard et al.2021.
A deeper understanding of the trophic role and interaction of wrasses, their predators and prey, as well as how the fisheries are affecting them, is a prerequisite for moving beyond speculations on the wider ecosystem responses of wrasse fisheries. The role of wrasses as cleaner fish in nature is also largely unknown. The cleaning behaviour of wrasses is likely to be opportunistic, since there are very few reports of such observations in natural conditions.[9]Breen 1996. Although it appears unlikely that feeding on parasites on other species is an important food source for Scandinavian wrasses, we cannot exclude the possibility that wrasses have an important ecological role for reducing parasite loads on other species.
The fishery could also impact the ecosystem if there is a high by-catch of other species resulting in catch-related mortality or if they are retained to be used for bait or human consumption. In Norway and Sweden, the regulations state that all bycatch species must be released on site immediately. The survival of by-catch is probably high since fishing gears are deployed in shallow waters and the traps must be hauled every day. In Sweden, an exemption from the landing obligation depending on high survival of species such as cod is a prerequisite for the fishery. In Norway, by-catch of crustaceans and fish constituted 9 % of the total catch in numbers of the reference fishermen in 2019-2020 (Figure 6). Fyke nets catch more by-catch than traps,[10]Halvorsen et al. 2017. but fyke nets are rarely used in Norway in recent years, and it is prohibited to use them for fishing wrasses in 2021. In the traps, cod (Gadus morhua) is the most frequently caught bycatch in Mid and Southern region, while green crab (Carcinus maenas) is more common in Western Norway. There has been concern from the general public regarding bycatch of the threatened European lobster (Homarus gammarus), but this species makes up only 0.15 % of the total catch in numbers and the majority of the lobsters were smaller than the minimum size limit of 25 cm (32 out of 38;[11]Halvorsen et al. 2020b.).
Footnotes
- ^ Costello 1991; Thangstad 1999; Skiftesvik et al. 2014.
- ^ Sayer 1999; Villegas-Ríos et al. 2013; Halvorsen et al. 2016b.
- ^ Alvsvåg 1993; Deady and Fives 1995a,b; Sayer et al. 1995a, 1996b, Bourlat et al. 2021.
- ^ Steven 1933; Rui Beja 1995; Nedreaas et al. 2008.
- ^ Alvsvåg 1993; Deady and Fives 1995b,a; Sayer et al. 1995b.
- ^ Eriksson et al. 2011.
- ^ Östman et al. 2016.
- ^ Dehnhard et al.2021.
- ^ Breen 1996.
- ^ Halvorsen et al. 2017.
- ^ Halvorsen et al. 2020b.
Figure 6. The most common bycatch in the Norwegian fishery visualized as the percentage of total catch (wrasses all sizes + bycatch in numbers) in each of the three management regions. Some of the species (e.g. Gobids); Gobiidae are difficult to identify precisely by non-experts and in those cases, we report the lowest taxonomic level that we are safe to assume correct identification (family or subfamily). Data from all reference fishers in 2019-2020 (N = 1535 pot hauls).
2.4 Translocation and escapement from aquaculture
Farmed fish escaping aquaculture is a serious threat to wild fish populations, through ecological interactions such as competition and predation, transfer of diseases and pathogens, and gene flow through interbreeding.[1]Jensen et al. 2010. There are many examples from open pen farming of salmonids, where escapees have hybridized with local river populations, leading to genetic swamping and reduced fitness.[2]Glover et al. 2017, 2020; Bolstad et al. 2017, Wringe et al. 2018. Consequently, Norwegian law states that aquacultures are obligated to report all escaping fish from aquaculture installations, but presently this only applies to farmed (salmonid) fish species, and escaping cleaner fish are left unreported.
Obviously, not only farmed fish pose a threat to wild populations. Transferring cleaner fish individuals between populations can equally affect native populations through disease transfer, and result in irreversible genetic changes. Such changes could involve shifts in allelic composition, loss of genetic variation, erosion of local adaptation and/or breakdown of population structure.[3]Laikre et al. 2010. IMRs risk assessment point to import and long-distance translocation of wild wrasse as important risk factors in terms of both disease transmission and changes in the genetic structure. Evaluation of the risk of translocation should not only include wrasse imported from Sweden but also the existing knowledge of genetically distinct populations within Norway. The lack of knowledge regarding the source and destination of cleaner fish transported within Norway is a big obstacle to assess and address the challenge of escapees.
In recent years we have gained an increasing knowledge of the overall population structure for all three wrasse species in Scandinavia.[4]Jansson et al. 2017, 2020; Faust et al. 2018, 2021, Blanco Gonzalez et al. 2016, Seljestad et al. 2020. Based on these studies it is clear that populations of these species are genetically structured along the Norwegian coastline. Even the species with the weakest structure, goldsinny wrasse, shows a general pattern of genetic isolation by geographic distance demonstrating that large scale translocations are likely to present an issue. The considerably stronger population divergence between Skagerrak and North Sea populations, as observed in both corkwing and Ballan wrasse, raises concerns of how translocation might alter this structure and affect native populations. The ongoing inadvertent translocation of wrasse via current aquaculture practice, from Sweden and southern Norway, to western, middle and North Norway, will likely result in mixing of fish from highly distinct genetic groups. While genetic change of local populations due to translocations has already been proven to take place for corkwing wrasse,[5]Faust et al. 2018. further studies are needed to investigate if, where, and to which extent translocated wrasse survive and hybridize with local wild populations for the other two species. There are indications from corkwing wrasse studies that hybridization takes place mainly in small, marginal and/or newly established populations in the northern edge of the species distribution.[6]Faust et al. 2021. It is possible, however, that the low number of escapees and hybrids detected further south along the western Norwegian coast could simply be a result of less translocated escapees, lower escapee survival or possibly a sampling effect since escapees may be difficult to detect in genetic surveys if local populations are large. Hence, it is important to implement better reporting and documentation of how cleaner fish are moved and when they escape. An obstacle for effective management is that the current practice of cleaner fish use is poorly documented and regulated.
Norwegian transporters are still not required to log and report the source, or the destination of wild-caught cleaner fish caught in Norway. Given that the largest population discontinuity is not aligned with country borders but can be found between Skagerrak and the North Sea, it is important to know the destination of wrasse caught in Norwegian Skagerrak. Not knowing which populations and what quantities are being used in different regions makes it difficult to assess and address the problem of escapees. Since 2017, all wrasse imported to Norway must be reported, detailing source, destination, and number of fish of each individual species. However, Swedish wrasse only constitutes 2-5% of all wild caught wrasse used in Norwegian aquaculture. There is presently no commercial fishing for wrasses in Denmark, and therefore no management plans in place.
The health status of the wild caught wrasses is poorly known. Repetitive transportations may increase the risk of introduction and spread of pathogens.[7]Peeler and Feist, 2011. Establishment of pathogens in new areas will depend on environmental factors and susceptible hosts. Some pathogens are adapted and specific to one or few hosts (specialists), while others easily adapt to new species and new environments (generalists). The current farm practice with its open design may contribute to transmission between farmed and wild fish by enabling pathogens to disperse to the surroundings freely / easily. In addition, the high density of hosts present in a farm situation might increase disease prevalence.[8]Peeler and Feist 2011. Even though there are no known examples with spread of disease through transportation of wild wrasse, there are several examples of pathogens being transported and introduced to new areas by transportation of live fish.[9]Egidius 1987, Gozlan et al. 2006, Peeler et al. 2011; Peeler and Feist 2011.
Conclusions and recommendations
We recommend that both fishery-dependent and -independent surveys are carried out to monitor the changes in species composition, abundance and distribution in the wrasse stocks and other species that can be directly or indirectly affected by the fishery. The challenges with highly variable CPUE should be carefully considered in survey design and choice of gear (Figure 3). A survey aiming to produce a stock indicator for wrasse should standardize gears and utilize data loggers to get associated environmental variables, such as temperature and depth.[10]Halvorsen et al. 2020a. In some areas, such as in Southern Denmark, salinity can also be a useful environmental variable to monitor when estimating abundance. Length data also has high value and allows length-based stock assessment models to be used, which may be appropriate for the wrasse species. Fishery-independent surveys conducted in and outside marine protected areas are effective for assessing the impact of wrasse fisheries given the high site fidelity of these species. Replicated study designs (at least three MPA – control sites, e.g.[11]Halvorsen et al. 2017.) and gathering of data on the fishing intensity in control sites (past and current) would be advantageous. Comparative studies including several sampling gear/methods is recommended to quantify bias, variability and can enable data from different surveys methods to be compared. Baited or un-baited remote video, or ROVs can be a promising alternative to catch-based surveys or snorkelling. Automated methods for identifying and counting wrasse and other coastal fish species in videos under development and can be used in the near future.[12]Knausgård et al. 2020.
Case studies have shown that the effects of fishing on local populations can be variable, ranging from no discernible effect to considerably affected. The fishery is species, size and sex selective, which can lead to unbalanced harvesting and reduced reproductive potential. Corkwing and ballan are the species with the highest overlap with the fishery and may therefore be more vulnerable to local depletions.[13]Halvorsen et al. 2020a. If wrasse fisheries change species composition or size structure, this may in turn affect functional ecology and trophic interactions. Wrasses are important mesopredators, but there are few published studies on the ecological links between wrasses and other ecosystem components that go beyond correlations and descriptions. We highlight the need for dedicated experiments to understand how wrasse populations are affected, specifically, to estimate fishing mortality linked to fishing effort, selectivity (species, size and sex) and to monitor wrasse populations routinely in and outside marine reserves, combined with data on spatio-temporal distribution of fishing.
Long distance translocation of cleaner fish between distant populations can affect native populations through disease transfer and result in irreversible genetic changes. Norwegian law states that aquacultures are obligated to report all escaping fish from aquaculture installations, this should also apply to escaping cleaner fish. Norwegian transporters are still not required to log and report the source, or the destination of wild-caught cleaner fish caught in Norway. Given that the largest population discontinuity is not aligned with country borders but can be found between Skagerrak and the North Sea, it is important to know the destination of wrasse caught in Norwegian Skagerrak. Not knowing which populations and what quantities are being used in different regions makes it difficult to assess and address the risk of escapees. In order to estimate the threat of translocation we need to know the source and destination of all wild caught cleaner fish, not just imported ones. If possible, long-distance translocation of wild cleaner fish should be avoided.
Footnotes
- ^ Jensen et al. 2010.
- ^ Glover et al. 2017, 2020; Bolstad et al. 2017, Wringe et al. 2018.
- ^ Laikre et al. 2010.
- ^ Jansson et al. 2017, 2020; Faust et al. 2018, 2021, Blanco Gonzalez et al. 2016, Seljestad et al. 2020.
- ^ Faust et al. 2018.
- ^ Faust et al. 2021.
- ^ Peeler and Feist, 2011.
- ^ Peeler and Feist 2011.
- ^ Egidius 1987, Gozlan et al. 2006, Peeler et al. 2011; Peeler and Feist 2011.
- ^ Halvorsen et al. 2020a.
- ^ Halvorsen et al. 2017.
- ^ Knausgård et al. 2020.
- ^ Halvorsen et al. 2020a.
3. Management considerations for sustainable fisheries for cleaner fish
This chapter describes different management measures that have been applied for wrasses, and we discuss the specific challenges that need to be carefully considered to ensure that the regulations are effective. Where relevant, we summarise how they have been approached and dealt with by scientific and management institutions in Scandinavia and the UK. We also briefly describe the fishery of lumpfish, which currently occurs at a much smaller scale compared to the wrasses, as lumpfish are only caught for broodstock.
3.1 Overview of management in each country
In Norway, wrasse fishing takes place all along the coast up to the southern border of Nordland county (halfway up the Norwegian coastline), and there are three geographically defined management units, which we refer to later in the text; South (Swedish border – Varnes lighthouse, Lista), West (Varnes lighthouse, Lista – 62 degrees North) and Mid (62 degrees North and northwards). See map in Appendix 1. The Norwegian fishery has primarily been managed by a mixture of minimum size limits, gear restrictions, restricted access to the fishery and in later years quotas on the regional and vessel level.
In 2016 a total quota of 18 million wrasses (not species specific) was implemented in the Norwegian regulations,[1]www.fiskeridir.no, Table 2. of which 4, 10 and 4 million were allocated to the Southern, Western and Northern management areas, respectively. The size of the quota was set similar to the catch level of 2015 in the southern and northern management areas and reduced by approximately 20% in the western area reflecting a similar reduction in catch-per-unit-effort (CPUE) for the reference fishers in the western area. Despite this, the fishery was not closed when the quotas were exceeded, and the Norwegian wrasse catches of wild wrasse peaked at around 28 million individuals in 2017. Stricter regulations were thereafter enforced, including a cap on the number of licenses. Consequently, the landings decreased to 18.5 million and 19 million wrasses in 2018 and 2019, respectively (Directorate of Fisheries). The Institute of Marine research reports the results of the stock assessment in October every year, and the changes in CPUE and length can be used to adjust the quota advice provided by IMR to the Directorate of Fisheries, which ultimately decide the quota for the coming season. The annual changes in CPUE and mean length from the former and the new survey have been used as supplementary data for scientific advice on quotas and size limits.
The fishery in Sweden is small relative to the Norwegian, but more than 800,000 wrasses were caught at the Swedish west coast in 2018 and sold to Norwegian farms. Catch quotas are not used in Sweden since there are no fishery independent measures of CPUE and size distribution to inform management. The management authorities have up to now decided to regulate effort by restricting the gear and the number of participants in the fishery (Table 2).
There are currently no management regulations implemented in Denmark as the fishery has not yet been established.
On the south coast of the UK, the fishery remained relatively unregulated until the relevant Inshore Fisheries Conservation Authorities (IFCAs) introduced regulations for each region as a coordinated management response.[2]Davies 2016, Street et al. 2017, Gravestock 2018. Each of the regional IFCAs developed exclusive, region-specific management responses from the offset. These management responses include minimum and maximum reference sizes (slot sizes) for retained wrasse, closed fishing seasons and areas where the fishing of wrasse is prohibited. Also included in some of the guidelines/byelaws are gear restrictions, maximum fishing depths, monitoring requirements for fishermen to fulfil, and recommendations for biosecurity and husbandry of wrasse.
Footnotes
- ^ www.fiskeridir.no, Table 2.
- ^ Davies 2016, Street et al. 2017, Gravestock 2018.
Table 2. Development of regulations in the Norwegian wrasse fishery since 2011.
| REGULATION | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | |
| Fishing opening date | |||||||||||
| South | May 30 | May 29 | May 27 | May 27 | June 10 | July 11/ July 61 | July 17 | July 17 | July 17 | July 17 | |
| West (south of 62°N) | June 20 | June 18 | June 17 | June 17 | July 1 | July 11/ July 6 | July 17 | July 17 | July 17 | July 17 | |
| Middle Norway (north of 62°N) | July 4 | July 2 | July 1 | July 1 | July 16 | July 251 | July 31 | July 31 | July 31 | July 31 | |
| Minimum size | |||||||||||
| General | 11 cm | ||||||||||
| Exception for Trøndelag and north | 9 cm | 10 cm | 10 cm | No exception | |||||||
| Corkwing-/Ballan wrasse | 12 cm/14 cm | ||||||||||
| Husbandry | Requirement for daily cleaning except for Sundays and public holidays | ||||||||||
| Fishing gear (commercial) | |||||||||||
| Number of fishing gear- from Varnes/ Lista and north | No gear limitation after fishing has been opened throughout the country | No gear limitation | 400 pos/fyke nets together all year | ||||||||
| Number of fishing gear- south | No gear limitation after fishing has been opened throughout the country | 100 pots/fyke nets together all year | |||||||||
| Gear | Pots and arrangement with exemption to use fyke nets for wrasse | ||||||||||
| Participation-recreational fishing | Fishing with pots and fyke nets is prohibited. Recreational fishers with a delivery agreement can apply for permission to fish. | ||||||||||
| Selection devices | No requirement for selection device | Requirements for entry barrier and escape openings | |||||||||
| Quota | |||||||||||
| South | No quota | 4 mill. | 4 mill. | 4 mill. | 4 mill. | 4 mill. | |||||
| West (south of 62°N) | No quota | 10 mill. | 10 mill. | 10 mill. | 10 mill. | 10 mill. | |||||
| Middle Norway (north of 62°N) | No quota | 4 mill. | 4 mill. | 4 mill. | 4 mill. | 4 mill. | |||||
| Closing date | |||||||||||
| South | No closing date | Sept. 2 | Dec 1 | Oct 1 | Oct 20 (Oct 232) | Oct 20 (Oct 232) | |||||
| West (south of 62°N) | No closing date | Sept. 9 (13/092) | Dec 31 | Oct 31 | Oct 20 (Oct 232) | Oct 20 (Oct 232) | |||||
| Middle Norway (north of 62°N) | No closing date | Sept. 23 27/092) | Dec 31 | Oct 31 | Oct 20 (Oct 232) | Oct 20 (Oct 232) | |||||
Table 3. Development of regulations in the Swedish wrasse fishery since 2011
| REGULATION | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 |
| Fishing opening date | ||||||||||
| May 15 | May 15 | May 15 | May 15 | May 15 | May 15 | May 15 | May 15 | May 15 | May 15 | |
| Size limits | ||||||||||
| Goldsinny | min 11 cm | |||||||||
| Corkwing | min 13 cm | |||||||||
| Ballan | 15 - 30 cm | |||||||||
| Husbandry | ||||||||||
| Requirement for daily cleaning | ||||||||||
| Required release | ||||||||||
| Spawning individuals, females with roe, undersized fish and all other by-catch must be released under water | ||||||||||
| Fishing gear (commercial) | ||||||||||
| Number of fishing gear | Limit of 50 pots / fyke nets per fisherman and 14 exemptions to be evaluated and renewed yearly to result in a maximum effort of 700 gear in the water at any one time | |||||||||
| Gear | Arrangement with exemption to use fyke nets and pots without escape openings for wrasse | |||||||||
| Participation-recreational fishing | ||||||||||
| No recreational fishery | ||||||||||
| Selection devices | ||||||||||
| No requirement for selection device, but has been tested during 2020 for inclusion in 2021 | ||||||||||
| Quota | ||||||||||
| Fishery not regulated by quota but by effort | ||||||||||
| Closing date | ||||||||||
| Dec. 31 | Dec. 31 | Dec. 31 | Dec. 31 | Dec. 31 | Dec. 31 | Dec. 31 | Dec. 31 | Oct. 31 | Oct. 31 | |
The lumpfish fishery and stock assessment
Wrasses stop being efficient de-lousing agents under a temperature of 6 ⁰C because of winter dormancy, and this has motivated interest for alternate cleaner fish species.[1]Powell et al. 2018b. Lumpfish (Cyclopterus lumpus), which are present across the Nordic and Arctic seas, are active and continue feeding at temperatures as low as 4 ⁰C.[2]Nytrø et al. 2014. Since, it is juvenile lumpfish that are effective as cleaner fish against salmon lice, these can be rapidly available (4 months) to deploy in salmon net pens.[3]Helland et al. 2014. These are reared in aquaculture from wild-caught adult lumpfish. While the traditional lumpfish fishery targeted only females for their roe, a smaller-scale fishery now targets mature females and males to be used as broodstock in the lumpfish farms to produce juveniles.
In Norway, the lumpfish roe fishery has been regulated since 1988, but only in the most northern counties (Nordland, Troms and Finnmark). Spawning starts in March and peaks in May. Since 2006, fishing is limited to the period before 20 June and before 5 July for East Finnmark where spawning occurs slightly later in the season. Anyone can participate in the fishery if their boat is below 13 m in length. The fishery is also regulated through a TAC (Total Allowable Catch) per boat. The minimum mesh size is 267 mm.
The Institute of Marine Research (IMR) has given advice for the lumpfish fishery in Norway since 1995. Stock assessment was based on commercial fishery data up until 2009. Since 2012, the advice is based on data from pelagic scientific surveys in the Barents Sea and the Norwegian Sea. A stock biomass index and abundance are calculated separately for juvenile and adult fish. Lumpfish abundance has increased substantially since the 1980’s and fluctuations are very much correlated to temperatures changes. Today, fishing mortality represents less than 1% of the standing stock of estimated spawners in the Barents Sea.
3.2 Management units
Intense fishing pressure raises concerns for potential overfishing, especially if population structure is not considered when defining management units. It is important that management units are based on meaningful biological entities, such as local adaptation and population structure. Otherwise, we risk overexploiting genetically distinct populations and removing local adaptation without the possibility of recovery.[1]Reiss et al. 2009, Funk et al. 2012. The wrasses and lumpfish are sourced from a large geographic area, which underscore the importance of mapping and accounting for genetic structure and population connectivity when defining the geographical units for fisheries management of these species. The three species of wrasses are known to have high site fidelity, limited depth range and small home ranges.[2]Sayer 1999, Villegas-Ríos et al. 2013, Halvorsen et al. 2016b, Halvorsen et al. 2020a. Thus, we would expect any connectivity between populations at a larger scale (> 10 km) to occur during the larval stage. This low level of migration would in turn lead to relatively high genetic differentiation between populations. This is true when comparing wrasse from the North Sea coast to the wrasse on the Skagerrak coast where populations differ, more or less depending on species.[3]Jansson et al. 2017, 2020, Faust et al. 2018, Seljestad et al. 2020. However, there is little to no genetic structure within these two areas, especially within the Skagerrak area which is genetically very homogeneous.[4]Jansson et al. 2017, Seljestad et al. 2020, Faust et al. in prep.
Considering the observed weak differentiation within the Skagerrak area, local overharvest or even extinction of local populations would likely result in only minor loss of the total genetic variation. This is only true, however, if the remaining effective population size is large enough to withstand the effects of genetic drift, i.e. if overexploitation does not occur in all local management areas simultaneously, and migration and gene flow among areas are still frequent. Recent studies in Skagerrak show higher abundances of wrasses within marine protected areas (MPAs) compared to outside.[5]Halvorsen et al. 2017. This would indicate that sub-populations depend more on local recruitment than immigration, and that it might take a long time for overfished areas to be recolonised from adjacent sub-populations. Hence, larval and egg dispersal should be taken into consideration when defining appropriate management units for the different species. When this knowledge is available, it should be effectively communicated to the relevant management authorities for a reconsideration of the spatial distribution of fishing effort and regulations.
Footnotes

The male cuckoo wrasse is one of the most beautiful fishes in the Nordic marine fauna.
3.3 Quotas
Quotas is one of many methods for regulating fishing effort but is currently only used in Norway where regional quotas are based on annual advice from the Institute of Marine research, but ultimately decided by the Directorate of Fisheries. However, each vessel with permit to catch wrasse has a guaranteed quota of 45-50000 wrasse, which have resulted in the regional quotas being overfished in western Norway and underfished in the Southern and Northern Region. s partly because wrasse permits can be sold across the management units, and there are currently no redistribution mechanisms that adjust the vessel quotas if there the numbers of vessels with permits in each region changes.
From a biological perspective, it is very challenging to provide advice for the size of the quotas in each of these large regions, since wrasses have very low mobility, which implies that there are a large number of demographically closed populations within each region. In other words, populations in the parts of a region where the fishing pressure is low are probably little affected by heavy exploitation happening elsewhere in the region. However, using trends in CPUE from the reference fishers (chapter 2.1), it can be possible to develop harvest control rules that can be used to adjust the quotas in the near future.
3.4 Size limits
Size limits are a common and widespread management tool for coastal fisheries. Scientifically, the guiding principle is to allow fish to spawn at least once before reaching harvestable size.[1]Froese 2004. However, several of the wrasse species have complex life histories and reproduction strategies, such as sequential hermaphroditism (ballan and cuckoo wrasse), alternative male life history strategies (corkwing and cuckoo) and differences in growth and maturity depending on the sex (corkwing, goldsinny). Thus, in these cases, the “minimum size limit” tool is unable to equally protect both sexes and male strategies. For ballan, cuckoo and corkwing wrasse, the males provide obligate parental care and are considerably larger than females. A reduction of the proportion of nesting males can have consequences for population productivity.[2]Kindsvater et al. 2020. Slot size limits or harvest slots, where both small (immature) and large (old, highly fecund) individuals are protected, can maintain or improve size/age structure and reproductive potential of the population.[3]Froese 2004, Hixon et al. 2014; Gwinn et al. 2015. Slot size limits has also been proposed to balance sex-selective fisheries for sequential hermaphrodites and sexual dimorphic species,[4]Hamilton et al. 2007, Halvorsen et al. 2016a, Kindsvater et al. 2020. and has been implemented for all five species in the Southern UK.[5]https://www.devonandsevernifca.gov.uk/content/download/2766/21147/version/2/file/Live+Wrasse+Fishery+Data+Analysis+November+2018.pdf.
In Norway, a general, non-species-specific minimum size limit of 11 cm was first implemented in 2011. Hence, the large differences in life history traits between species (including size at maturity) was not accounted for.[6]Potts 1974, Dipper et al. 1977, Dipper and Pullin 1979, Costello 1991, Darwall et al. 1992, Sayer et al. 1995b, 1996b. Later, in 2015, the size limits for ballan and corkwing were raised to 14 cm and 12 cm, respectively, but this has not been shown to reflect size at maturity, especially for Ballan. IMR have suggested that size limits are revised and are set according to the life history for each species.[7]Table 7, Halvorsen et al. 2020b. In Sweden, the minimum size limits in Sweden are more conservative compared to Norway. Goldsinny has the same size limit (11 cm), corkwing slightly larger (13 cm) and ballan has both a minimum and maximum size (15-30). In the UK, IFCAs on the south coast also introduced minimum and maximum size limits (slot sizes) in 2017.These sizes varied between IFCA districts, were species-specific and based on evidence from the literature on size at maturity along with negotiations with salmon farmers and fishers. The minimum size for goldsinny and rock cook was 12 cm in all three districts, with maximum size limits likely being biologically arbitrary at 22 cm, 23 cm and 18 cm in Cornwall, Devon & Severn and Southern districts, respectively. Corkwing wrasse in Cornwall and Devon & Severn’s districts were subject to the same size restrictions as goldsinny and rock cook in 2017, but in the Southern district corkwing minimum and maximum size limits are 14 cm and 22 cm. Following Devon & Severn IFCA’s first analysis of fishery dependent on-board observer survey data, which highlighted that 94% of corkwing wrasse in the district were being retained once caught, the minimum and maximum size limits were amended to 14 cm and 18 cm in 2018.[8]Curtin et al. 2017. Ballan wrasse minimum and maximum size limits also vary between districts; Cornwall: 16 and 18 cm, Devon & Severn: 15 and 23 cm, Southern: 18 and 28 cm. Following the introduction of Cornwall IFCAs limited permit byelaw in 2019, maximum size limits for all wrasse species were removed. In 2017, when the management measures were introduced, cuckoo wrasse had the same slot size limits as Ballan wrasse in the Cornwall and Devon & Severn districts. Cuckoo were not permitted to be retained on-board in the Southern district, and since the introduction of the byelaw in Cornwall the retention of cuckoo was prohibited.
Footnotes
- ^ Froese 2004.
- ^ Kindsvater et al. 2020.
- ^ Froese 2004, Hixon et al. 2014; Gwinn et al. 2015.
- ^ Hamilton et al. 2007, Halvorsen et al. 2016a, Kindsvater et al. 2020.
- ^ https://www.devonandsevernifca.gov.uk/content/download/2766/21147/version/2/file/Live+Wrasse+Fishery+Data+Analysis+November+2018.pdf.
- ^ Potts 1974, Dipper et al. 1977, Dipper and Pullin 1979, Costello 1991, Darwall et al. 1992, Sayer et al. 1995b, 1996b.
- ^ Table 7, Halvorsen et al. 2020b.
- ^ Curtin et al. 2017.
| Species | Parental care | Sex change | Max age – length | Length at maturity | Min size limit | Advised --Min size (–> max size) |
| Ballan | Yes | Yes | 29 yrs – 50 cm | Fem: 22 cm. Males: 30-40 cm | 14 cm | 22 (–> 28) cm |
| Corkwing | Yes | No | 9 yrs – 25 cm | Females: 10 cm, N. Males: 15 cm, Sneaker males: 8 cm | 12 cm | 13 cm |
| Goldsinny | No | No | 20 yrs – 20 cm | 8 cm | 11 cm | 11 (->14) cm or 12 cm |
Table 7: Summary of key reproductive and life history traits of the five wrasse species.
Life history data is based on both published and unpublished data (from yearly surveys) and is here representing western Norway mainly. Colours indicate whether current size regulations are in line with reproduction/life history in each species; red: size limits not reflecting biology, yellow: some reflection of biology and green: size limits set in good accordance with biology. N. males = Nesting males.
3.5 Gear regulations
Gear regulations are also common and include measures regulating the type of gear to be used, the maximum entrance size on traps and fyke nets, and the size and position of escape openings. Even subtle alterations in gear design can affect size selectivity and catch efficiency for both target species and bycatch.
When the Norwegian fishery started in the 1990’s, fyke nets for catching eels were the predominant gear used, but the proportion of fyke nets to traps decreased from 78% to 14% from 2008 to 2009.In 2018, only 1% of the reported Norwegian wrasses catches were taken by fyke nets.[1]Directorate of Fisheries, sales-notes statistics. To prevent bycatch of otters entry openings were restricted and to reduce catches of undersized wrasse, escape openings became part of the national technical gear regulations from 2015.[2]Table 2, Jørgensen et al. 2017. In 2011 most fishers used gear without escape panels (for the smallest fish to escape), but gradually the fishers saw the benefit of not catching undersized fish, and hence experimented themselves with different escape panels. It was not until 2015 that it became mandatory to use escape panels with minimum grid openings of 12 mm. For the Skagerrak coast there is a gear limit of 100 units (combined number of pots and fyke nets) per vessel, while the limit is 400 units for the remainder of the coastline. The gear should be tended daily, except Sundays. Fyke nets will be prohibited from 2022 and the entrances in traps must be circular with a maximum diameter of 6 cm.
In Sweden, there have been stricter regulations on the number of fishers and number of gears to be used, Currently, only 14 fishermen have been granted permission to fish wrasse along the Swedish west coast and with no more than 50 gear each. Different gear types are used for the different species, with a gradual reduction in the importance of fyke nets. Corkwing and ballan wrasse are mainly caught in fyke nets whilst pots are more efficient in catching goldsinny wrasse. Currently there are no regulations that call for escape openings or entry barriers, although some fishermen buying Norwegian pots keep and use the selection devices already in place. In 2019 SLU conducted a test of size selective gear for the Swedish wrasse fishery and found that escapes panels of 12x70 mm along the end of the pots and fykes nets were most efficient in preventing the catches of undersized fish. For 2020 the fishermen have been commissioned to test selection gears so that their use can be worked out in the 2021 permits.
Since the fishery began on the south coast of the UK, fishers in all three districts predominantly fished with pots. Cornwall IFCA have a permit condition within their byelaw that pot traps cannot exceed length 75 cm, width 45 cm, depth 35 cm (including exterior attachments to the trap). Pots in all districts must include a hard/rigid eye that does not exceed diameter 9 cm and at least two unobstructed rigid vertical escape gaps (7 cm height, 1 cm width). There is no limit to the number of pots a fisher can have in Cornwall, but in Devon & Severn and Southern districts there are pot limits of 120 and 80 pots per vessel, respectively. Early in the development of the fishery, some fishers in the Southern district began targeting only Ballan wrasse using rod and line. Now a majority of the active wrasse fishing vessels in the Southern district fish using rod and line (either exclusively or alongside the use of pots). In the other districts there has only been limited use of rod and line to target wrasse. Ballan wrasse are the most sought-after species on the south coast, being sold to salmon producers in some areas for over £17 per fish, which has driven the change towards a more targeted method of fishing.
3.6 Bycatch regulations
Most fisheries want to minimize catch of non-target species, and there are several measures that can be used to achieve this in the wrasse fisheries, either through gear modifications to reduce catch rates or specific regulations regarding the handling of bycatch when captured.
In both Norway, Sweden and UK, it is compulsory that all non-target species must be released immediately at the site of capture. The fishery takes place in shallow waters (<6 m), which minimizes the risk of barotrauma. Based on the experience in the scientific fisheries, it is reasonable to assume that most fish and crustaceans survive being captured and released in good condition, given that the gear is hauled at least once per day and immediate release in shallow water. The most frequently caught by-catch species are cod and green crab (Figure 6). In addition, there were occasional reports of bycatch of seabirds and otters in fyke nets. IMR has monitored bycatch from a selection of fishermen along the Norwegian coast, which has revealed considerable geographical variation. In Sweden, fishers must also release ready-to-spawn and injured/sick individuals. Only bycatch of cod, eel and lobster are reported by Swedish fishers. Survival of released species are unknown but assumed high for quota species in the exemption from the landing obligation.
In UK, the Cornwall IFCA record species that are caught as bycatch during their on-board observer surveys. Cuckoo wrasse in the Cornwall district and Southern district are required to be returned to the sea immediately if caught.
3.7 Closed seasons
The reproductive behaviour of wrasses is extraordinarily diverse among the five species; but the common denominator is a spawning period in May – July where males are territorial. The species with parental care (corkwing, ballan, cuckoo) are especially vulnerable for fishing during this period.[1]Halvorsen et al. 2016a, Kindsvater et al. 2020. The onset and duration of the spawning period seem to be influenced by temperature but appears to be similar in Western and Southern Norway.[2]Halvorsen, Skiftesvik et al. unpublished data.
Figure 8 illustrates the spawning time for corkwing wrasse, and when the fishery opened in Norway in the different years. The fishery in Sweden opens the 15th of May, and there are no restrictions on fishing period in Denmark. On the south coast of the UK, closed seasons vary between IFCA districts. In 2017 in the Southern and Devon & Severn districts the fishery was closed between 1st April and 30th June, but in Devon & Severn this was changed in 2018 to 1st May to 15th July based on records of spawning individuals seen during observer surveys in 2017. When Cornwall first introduced management measures, closed seasons were introduced between 1st December and 28th February, with a second closed season between 1st April and 30th June, in the area adjacent to Devon & Severn’s district. The first closed season was then changed to between 1st January and 31st March with the introduction of the byelaw as a result of a spawning study completed further east on the south coast (unpublished data). Closed seasons were implemented based on evidence of spawning seasons from the literature, and with negotiation with fishers and salmon farms. There is a need for more localised studies that identify the spawning seasons for wrasse on the south coast of the UK.

Figure 8: The proportion of spawning corkwing as a function of week number 2013-2017 for corkwing wrasse, illustrated with a modelled smoother. Dots represent yearly observed data, while vertical lines show the opening date of the fishery in each year.[1]Halvorsen, Skiftesvik unpublished data.
Footnotes
- ^ Halvorsen, Skiftesvik unpublished data.
3.8. Closed areas
Marine protected areas are effective tools for management and conservation of sedentary species.[1]Baskett and Barnett 2015, Sørdalen et al. 2018, 2020; Moland et al. 2021. Given the low mobility of wrasses, even small (<1km2) marine protected areas (MPAs) can have positive effects on abundance, mortality, and size structure.[2]Halvorsen et al. 2016b, Halvorsen et al. 2017, Halvorsen et al. 2021. The abundance, species composition and size structure of wrasses is strongly shaped by the combination of wave exposure and depth,[3]Halvorsen et al. 2020a. knowledge which should be considered when designing MPAs with the purpose of protecting wrasses. Such an assessment was recently conducted for deciding optimal placement of a wrasse MPA in mid-Norway.[4]Kleiven et al., 2021. Norway has established 51 lobster reserves where stationary gears (e.g. traps, fyke nets, and in most of the areas also gillnets) are prohibited, thus no commercial wrasse fishing can take place.[5]Moland et al. 2021. Sweden has several marine protected areas where no wrasse fishing can take place. A survey conducted in 2016 comparing a 2.6 km² large marine protected area with two equally unprotected areas. Data showed no clear evidence of depletion for any of the three species.[6]Bourlat et al. 2021. However, very few ballan wrasse were caught in both areas making it difficult to evaluate differences. It has been suggested to close a fraction of the Natura 2000 areas in the Swedish west coast archipelago from wrasse fishery as reference areas. Early on the Swedish County Board took another direction, instead limiting the number of fishermen allowed in each Natura 2000 area. In the UK, each IFCA district on the south coast has introduced a range of small voluntary closed areas with their initial management measures. These have remained voluntary, with good compliance from fishers.
Recommendations
Due to the high spatial differentiation of wrasse populations relative to the current management units, it is difficult to set quotas that ensure sustainable fishing levels. Regional quotas may not be able to prevent local overfishing where the concentration of vessels is high. However, quotas provide an opportunity to limit and adjust overall fishing effort and used in combination of other technical regulations (size limits, gear restricts, closed areas) that reduces the likelihood of local overfishing, it can be useful ensure a predictable and sustainable supply of cleaner fish to the industry. With stricter regulations on size limits and the number of permits and gears, as is the case in Sweden and UK, quotas may not be necessary, and the catch will fluctuate with the natural variations in stock size.
It is recommended to exploit the full range of technical measures for wrasse management. Size limits should reflect each species life history. Although there is available growth and maturity data for most of the species, the spatial coverage is low, and should be improved in order to evaluate what spatial scales should be considered in management, especially in setting size limits. With data on life history and catchability, one may construct population models which explore the response to varying levels of fishing effort, selectivity (size limits, gear selectivity). This could be an important tool for providing advice for controlling fishing effort and optimizing size regulations, as well as predicting potential changes in absolute and relative abundance under different fishing and management scenarios. With such a framework, uncertainties can be accounted for and the different mechanisms determining population dynamics are jointly considered.
To reduce bycatch and the catch of large ballan, the entrance of the gear should be as small as possible (oval openings, 7.5 x 6 cm or circular 6 cm diameter). To allow small wrasse to escape, escape grids should be used. Bycatch and smaller wrasse should be immediately released at the point of capture, ideally through a pipe that allows the fish to be released below the surface to avoid predation by seabirds. Bycatch of other species constitute 10–20% of the catch in the Norwegian fishery. Similar assessment of bycatch should be done in other countries with active wrasse fisheries. The Norwegian regulations state that bycatch must be carefully released at the site of capture. This is not always the case, and it is advocated that the fishers’ behaviour in handling bycatch is further studied. Also, studies investigating post-release survival of key by-catch species under different release scenarios is needed.
Fishing should not be permitted during the spawning period. The onset and duration of the spawning period may vary geographically and between years, so it is advised to conduct a weekly scientific survey to assess this or to use a conservative approach (setting the opening date mid-July). Closed areas are efficient in preserving natural species composition and size structure and is viewed as a positive supplement to other technical management measures and has particularly high value for scientific surveys monitoring the impact of fishing.
4. Use of wild caught and farmed cleaner fish
The number of cleaner fish stocked in salmon and trout sea cages has increased from less than five million in 2009 to more than 51 million in 2020. (Source: Directorate of Fisheries) As described in section 1, “cleaner fish” refer to lumpfish and several species of wrasse (ballan wrasse, corkwing wrasse, cuckoo wrasse, goldsinny wrasse, rock cook). Until recently, cleaner fish have been almost exclusively wild caught, whereas the increased production of lumpfish outnumbers wild caught wrasses and amounts to around 34 million of the 51 million cleaner fish that are stocked in salmon farms. In the northern part of Norway, lumpfish is the only cleaner fish used since the temperature is too low for wrasses. Lumpfish and wrasse are both used further south. The main area wrasse is used is on the west coast of Norway. Advice around the use of wrasse and lumpfish in salmonid aquaculture can be found in www.rensefiskskolen.no. This site is updated with new knowledge as it becomes available.
The Norwegian Seafood Research Fund (FHF) and The Research Council of Norway (NFR) supported pioneering projects that aimed to increase knowledge of wild wrasses, how to breed ballan wrasse, and later also lumpfish, and how to best use cleaner fish in sea cages. They also funded projects aiming at understanding what causes loss of cleaner fish in sea cages, projects on feed development, vaccine development, and much more (see Table 8). Several projects are underway to find ways to improve knowledge on the welfare and mortality of cleaner fish, as well as determine the types and locations of facilities and seasons that are most effective for each species. Additionally, there are projects investigating feeding methods and strategies, as well as methods for sustainably fishing the various cleaner fish species, including reducing bycatch. A rapid increase in knowledge of these topics is expected in near future.
Some of the aspects that are currently being investigated are: mortality in sea cages (various causes) and in production, feed quality and feeding strategies, providing a suitable environment, correct use of cleaner fish species with respect to place and time, strategy for fishing out cleaner fish before handling of salmon that can affect cleaner fish, and use / reuse of cleaner fish. These aspects will be presented in this chapter.
4.1. Aquaculture production of cleaner fish
The first trials using wrasse as cleaner fish were performed with wild-caught fish. Wild-caught wrasses are still extensively used, but the idea of using farmed wrasse was adopted early. Attempts to farm several of the wrasse species were made in the early 1990’s.[1]Skiftesvik et al. 1996, Stone 1996. Ballan wrasse quickly emerged as the best candidate for farming. Later, lumpfish was introduced as a possible lice picker.
Ballan wrasse
It was found early on that farmed ballan wrasse was at least as effective at eating lice as wild-caught ballan wrasse.[2]Skiftesvik et al. 2013. The first farmed ballan wrasse came in 1996, but it was not until 2010- 2011 that companies tried to breed this species commercially.
Ballan wrasse has proven quite difficult to rear. Despite great effort in several farming companies, the production has not increased significantly. Only about 4 % of the more than 17 million wrasses used in 2019 were farmed ballan wrasse, increasing to 10 % in 2020 (source: Directorate of Fisheries). Several on-going projects are working with issues related to the breeding of ballan wrasse, but the industry will probably have to depend on wild-caught ballan wrasse and other species of wrasse for many years. However, an increased effort by many companies to farm ballan wrasse this year might increase the production of farmed ballan wrasse.
The farmers experiences so far with breeding of ballan wrasse were summarized by Amundsen and Størkersen (2019), but the problems in the ballan wrasse hatcheries have never been properly reviewed. A part of a new FHF project[3]project 39, table 8. is to do just that.
Most companies that started up with ballan wrasse farming early on, gave up after a few years of unsuccessful attempts. The first protocols for breeding ballan wrasse were developed over 10 years ago,[4]project 6, Table 8. and have been refined with new knowledge about the species over time. Previous projects[5]project 2 and 6, Table 8. found that the best breeding temperature regarding growth and survival was 14–16 °C. Problems related to spawning and collection of the sticky eggs were solved by using spawning mats.[6]project 7 in Table 8. Mats with fertilized eggs can be hung in incubators until the hatching is over. Attempts have been made to dissolve the sticky film to have eggs that do not stick, but these methods did not work well, and spawning mats are used today. The ballan wrasse's sexual maturation and gender change were also mapped in project 7, Table 8.[7]Muncaster et al. 2010, 2013. Project 2, Table 8, characterized the spectral sensitivity of larval-juvenile and adult Ballan wrasse, and based on that, recommended species-specific lighting for indoor intensive culture environments.[8]Loew 2012. The ballan wrasse’s genome was mapped in project 4, Table 8, and is archived here: http://www.ebi.ac.uk/ena/data/view/FKLU01000001-FKLU01013466
Lumpfish
The use of lumpfish has gradually increased as traditional methods to remove the salmon lice became less effective. Almost all lumpfish used as cleaner fish are farmed. There has been a sharp increase in the production in recent years, and in 2019 more than 42 million lumpfish were produced in hatcheries, and 42 million reported set out in salmon cages that year. In 2020, 32 million farmed lumpfish were reported to have been set out in salmon cages (source: Directorate of Fisheries).
As lumpfish tolerate lower temperatures than wrasse, the implementation was boosted in the northern parts of Norway. In the beginning, the production of juveniles was solely based on wild-caught brood fish, where eggs were stripped, incubated, hatched and reared to suitable size for transfer to commercial cages. The increasing demand for lumpfish has resulted in gradually moving from wild caught fish towards intensive cultivation. Lumpfish production is currently utilizing similar technology and techniques as those used in halibut, wrasse and cod aquaculture, and a breeding program for lumpfish was established in 2017. In 2020 a little over 32 million lumpfish were transferred to cages at commercial marine growth sites for salmonids, which constituted 63 % of all cleaner fish used that year.[9]Statistics from the Directorates of Fisheries. Therefore, lumpfish is the single most important cleaner fish species.
There are severe disease problems with lumpfish, and therefore there is an urgent need to determine potential health issues. In addition, many hatcheries have their own production management strategies. Feeding strategies vary widely between hatcheries with some employing constant feeding regimes whilst others use more restrictive regimes based on natural daylight strategies. There is no clear standardized approach to specific photoperiod lengths during the tank rearing phase.[10]Jonassen et al. 2018.
Two ongoing projects[11]8 and 9, Table 8. will develop protocols for keeping farmed brood stock and controlled reproduction, which is a prerequisite for achieving genetically based improvements.[12]10, Table 8. Protocols will be developed for the entire life cycle of lumpfish through systematic work with optimal brood stock farming, optimal fry farming and efficient use of lumpfish in cages with salmon.
Lumpfish reared in hatcheries have varying incidences of cataracts. Some hatcheries have suggested that up to 45% of their fish have developed cataracts just prior to transfer to sea cages. These fish will have limited visual acuity, and this may well be the reason why mortalities are observed a few weeks after transfer. If 40% of the lumpfish transferred to sea cages are visually impaired, then they are unlikely to graze on attached sea lice and the remaining fish may be unable to maintain a low sea lice burden in the cage. In future, more complete reporting of lumpfish mortalities and assessment of health status within hatcheries would identify favourable and unfavourable husbandry conditions and thereby lead to improvements. Amundsen and Størkersen (2019) summarized the farmers experience with rearing lumpfish. They experienced problems due to hygiene and water quality, “low quality” eggs and larvae and high mortalities of larvae and juveniles, typically with peaks in mortality in connection with the first feeding and after vaccination. They also report welfare problems such as tail biting, fin erosion and fin rot. Mortality was due to various bacterial and viral diseases.
Footnotes
- ^ Skiftesvik et al. 1996, Stone 1996.
- ^ Skiftesvik et al. 2013.
- ^ project 39, table 8.
- ^ project 6, Table 8.
- ^ project 2 and 6, Table 8.
- ^ project 7 in Table 8.
- ^ Muncaster et al. 2010, 2013.
- ^ Loew 2012.
- ^ Statistics from the Directorates of Fisheries.
- ^ Jonassen et al. 2018.
- ^ 8 and 9, Table 8.
- ^ 10, Table 8.
4.2. Transport of hatchery produced Ballan wrasse and lumpfish
A considerable proportion of cleaner fish are transported several hundred kilometres by road to their destined farm. Project 16 and 17, Table 8 investigated transport stress in cleaner fish. Handling in connection with transport is a stressor for lumpfish, while longer transport provides the fish with an opportunity to "stress down". The biggest challenge for commercial transport is reloading from primary transport (car) to secondary transport combined with the secondary transport often having an unstable water environment and being too short to allow time for recovery. After the secondary transport, a significant stress level has accumulated in the period just before the fish is transferred to sea cages. One aquaculture company has solved the problem with two transports - they drive the transport car on to a ferry and load the fish from the car into the sea cage directly. This way they have eliminated the secondary transport, and the company reported a much better lumpfish survival after this method was implemented. High stress levels when introduced into sea cages can increase the risk of developing chronic stress, leading to impaired immune responses which increases susceptibility to pathogens. Increased mortality is seen especially at sites with periods of strong currents.[1]Jonassen et al. 2019. For Ballan wrasse (farmed), sites with strong current and high temperature (21 °C) led to mortality after transport stress. Ballan wrasse are not stressed by high light intensity, they are tolerant of low salinity, and had no problem with temperature fluctuations of 3–4 °C.
Footnotes
- ^ Jonassen et al. 2019.
4.3. Size and transfer time for lumpfish
The size of lumpfish at which they are transferred to sea cages varies between commercial farmers. Some transfer fish as small as 20 g whilst others transfer at a minimum of 50 g. Some farmers still deploy lumpfish during winter months but more and more avoid winter deployment. A previous study by Imsland et al. (2016a), showed that small lumpfish (mean weight 20 g) deployed in winter had lower growth rates compared to larger fish for a period, after which growth rates increased. This reduced growth potential may be explained by low water temperatures and the fish required a period of acclimation after transfer to adapt to the different environmental conditions. The challenge in using lumpfish as biological de-lousing agents is to optimize their grazing potential. Small lumpfish do graze on attached sea lice from Atlantic salmon.[1]Imsland et al. 2014a. However, if lumpfish are subjected to disturbance that results in increased stress, then there is a higher probability that they will become prone to disease, particularly bacterial agents. More research is required to clarify best practices when lumpfish can be deployed in commercial salmon cages.
Footnotes
- ^ Imsland et al. 2014a.
4.4. Use of cleaner fish in sea cages with salmon
How well do they work?
There are few studies that quantify the efficiency of de-lousing, as the documentation is mainly based on observations, registrations and counts without reference to a situation without cleaner fish – meaning “without a scientific control”. In a recent paper, Overton et al. (2020) looked at the documentation of the cleaning effect in reports and publications and concluded that the effect of cleaner fish was poorly documented:
“Only 11 studies compared lice removal between tanks or cages with and without cleaner fish using a replicated experimental design. Most studies had insufficient replication (1 or 2 replicates) and were conducted in small-scale tanks or cages, which does not reflect the large volume and deep cages in which they are deployed commercially. Reported efficacies varied across species and experimental scale: from a 28% increase to a 100% reduction in lice numbers when cleaner fish were used”.
This review also revealed that the interaction of cleaner fish and salmon in commercial scale sea cages had rarely been documented. To compensate for this, Barret et al. (2020) performed a national scale analysis of data reported to the Norwegian Food Authorities by the farmers about cleaner fish use and lice levels during 2016-2018. This analysis only found weak and short-lived effects, and that most sites eventually had to resort to mechanical, thermal, or chemical de-lousing operations. They concluded that although some farms consistently obtained good results, there was a widespread sub-optimal use of cleaner fish. There is, however, continuous advancements in the husbandry and management of cleaner fish (e.g. Imsland et al. 2019a-b; 2020), and recent analysis of updated data suggests improved effects.[1]LT Barrett, pers. com. An important finding in Barret et al. (2020) is the farms that consistently obtained good results. It is crucial to study what they do right, and the findings here are shared with other farmers.
The studies by Overton et al. (2020) and Barret et al. (2020) are limited in that they only had available relatively coarse publicly available data[2]barentswatch.no. about sea lice levels and cleaner fish deployment at farms. Studies should be made on detailed, multi-factorial, industry data (e.g. Imsland et al. 2018) to get clear recommendations on which farms, seasons, fish sizes and rearing practices (shelters, feeding, etc) the different cleaner fish species work well.
Factors that influence the efficiency of de-lousing
Project 18, Table 8, and a follow-up experiment, documented lice-picking activity of lumpfish under full-scale production conditions. They also assessed how lice grazing was affected by season, density, and size of salmon. The proportion of lumpfish was either 3.75 or 7.50 % of the number of salmon in the sea cage. There were problems with high mortality, but nevertheless, lice infestation was reduced by approximately 50% in the group with 3.75 % lumpfish.[3]Imsland et al. 2018. From the summary and main findings: “There can be great variation in how efficiently lumpfish eat lice and the proportion of fish that eat lice can in some cases be low. In some cases, this is related to the fact that there are other types of food for the lumpfish that it prefers when it is easily available. Lumpfish is considered to be an opportunistic feeder, which means that they eat most things, but make choices (have preferences) if they have choices.”
In some experiments, it was observed that copepods were preferred as the main food when abundant. In other cases, the lumpfish fed exclusively on jellyfish. They found that flushing of the nets had negative effect of the lumpfish’s effectiveness as cleaner fish. The fish near the net wall suffered wounds and mucus loss which led to high mortality. They also found that small lumpfish are more effective than large lumpfish, and that lumpfish are active during daytime.
Combination of different cleaner fish species
Project 19, Table 8, tested combinations of the cleaner fish species Ballan wrasse, goldsinny, corkwing and lumpfish. The trials lasted only three weeks, but cleaner fish had clear effects on the number of salmon lice compared to controls. A short summary from the project report: In the treatments with each species individually compared with the control cages without cleaner fish, there were significantly lower lice numbers for all three wrasse species, but not for lumpfish. Ballan wrasse + goldsinny and Ballan wrasse + corkwing wrasse gave the best effect. The combination of goldsinny and Ballan wrasse also had significantly lower lice numbers compared to when these species were used individually. However, lumpfish stayed mostly at shallow depths, whereas wrasses and the salmon were more frequently observed in deeper positions during the experiment. Lumpfish was observed to feed on jellyfish, which there was a lot of, which might explain why they were less effective. Most goldsinny were seen at the lower end of the shelters. In combination with lumpfish, several cases were seen of goldsinny picking on salmon. In the combination with Ballan wrasse, it was still goldsinny that dominated in the lower areas of the sea cage, and here one could see some cases of lice eating by both goldsinny and Ballan wrasse. Corkwing wrasse generally seem to have somewhat more active behaviour than other wrasses. Some corkwing males were aggressive towards individuals of their own kind. This happened close to the shelters. This behaviour was not recorded in any of the other species. Ballan wrasses were rarely observed in the upper part of the cages. Further down, the ballan wrasse was commonly observed outside, among the salmon, and inside the shelters. In combination with goldsinny, there were more observations of Ballan wrasse inside the shelters than outside. Although this experiment with combinations of different wrasse species indicated a good effect on reducing lice on salmon and that cleaner fish works well together, there is still a need for a better understanding of seasonal variations in behaviour and lice grazing to get an optimal combination of species throughout the year.
Developing suitable shelters and feeding stations
Both wrasses and lumpfish need shelters and places to hide and rest. To obtain a better effect of the use of cleaner fish and a better survival, the fish farmers have developed solutions that provide shelter and feeding stations. Best practice manuals and guidelines were developed, based on scientific information and experience (e.g. Lusedata, URL: http://lusedata.no/for-naeringen/veiledere-leppefisk, and www.rensefiskskolen.no). It is important to remember that “cleaner fish” consist of several species with different preferences and needs. Through behavioral studies and video filming, they found species differences in relation to the use of different shelters in the winter when they had a choice. Wrasses preferred “Kinatare” while the lumpfish used hard plastic shelters.[4]Espmark et al. 2020.
Juvenile lumpfish are typically found among kelp during the first year of their life, both attached and free floating.[5]Ingólfsson and Kristjánsson 2002. Mimicking their natural requirements for surface adhesion it is reasonable to expect that lumpfish stocked in commercial salmon cages need some type of shelter or substrate to attach to when resting.[6]Imsland et al. 2015b, 2018b. Substrates may also provide shelter for lumpfish during periods of inactivity and/or extreme environmental conditions. There are several companies manufacturing a range of substrates for this species. Most resemble artificial seaweed (kelp) and are composed of PVC. These “kelp curtains” are normally deployed using a series of buoys to maintain a certain depth. One critical factor when deploying these artificial substrates is to ensure that sufficient surface area is available to the lumpfish, particularly as they grow. Irrespective of the type of substrate offered, it is critical to the welfare of the fish that they have access to some form of substrate due to the need to rest. To date, there has been no systematic review/research to determine if current shelters and curtain kelps are optimal for lumpfish. Shelters for lumpfish must be easy to remove/clean and lumpfish should not be injured when the shelters are removed. In addition, shelters must provide farmers visual access to observe behaviour of the fish.
All wrasses live close to seaweeds, kelps, and stones in their natural environment where they find their food, have shelters and resting places. They always rest at nighttime, and they reduce activity and can also rest through day and night in the coldest and darkest time of the winter. Shelters are a necessity when wrasses are used in salmon farms. As noted above, there is a range of products available. More tests are needed to find the best ones for summer and winter use. Feeding of wrasses should be done near shelters.
Lumpfish in combination with lice skirts used on salmon farms
Lice skirts are used to avoid the influx of pelagic salmon lice larvae (nauplii) in salmon farms. Lice skirts were not considered beneficial for lumpfish, but there is probably a great potential in optimizing such an operating strategy. Project 30, Table 8, is currently assessing the combined effect of skirts and cleaner fish, including the consequences for cleaner fish welfare. The project will identify main problems and improvement measures. Lice skirts have now been in use for almost 10 years, and a systematic review of their effect will reveal optimal use of this method under different environmental conditions, with or without cleaner fish. Another ongoing project (project 20, Table 8) working with optimization of lumpfish use, will test a user regime of cleaner fish that includes reducing the number of individuals, facilitating good health throughout the production cycle and streamlining each individual's capacity as a "lice eater". Farmed lumpfish will be transported from production facilities out at sea to salmon farms and they will investigate how lumpfish are affected by such transitions, both in terms of stressors and individual cognitive capacity to adapt to a new environment (se also 5.3).
Freshwater treatment (Louse or AGD treatment)
If salmon must be treated for lice or amoebic gill disease (AGD) with freshwater, it is important to know that neither ballan wrasse, corkwing nor goldsinny tolerate direct transfer to freshwater, but they tolerate two hours in freshwater if the salinity is gradually reduced first.[7]project 19, Table 8. The Norwegian “Akvakulturforskriften” states that «Før det utføres operasjoner på anlegget som kan føre til belastning på rensefisken, skal rensefisk sorteres ut og vernes mot skade og unødvendig påkjenning. Etter samråd med veterinær/fiskehelsepersonell, kan rensefisk bli stående i produksjonsenheten dersom dette tar bedre hensyn til fiskevelferden». When it comes to freshwater treatment, wrasses have to be separated from the salmon before freshwater treatment starts unless the freshwater is gradually introduced.
Feed development and feed requirements of wrasses
Feed and feed requirements emerged early on as a problem area for Ballan wrasse rearing. It was difficult to get the ballan wrasse to change from live feed to formulated feed. There were also often problems with survival of fish fed formulated feed, which had to do with digestive problems and lack of knowledge about the ballan wrasses' nutritional needs.
Many projects have investigated these issues. Project 5, Table 8, examined the digestive system with several approaches, and methods ranging from histology to next-generation deep sequencing. It was discovered that digestion of protein, fat and glycogen is very efficient, and that ballan wrasse have an efficient digestive system where up to 70 percent protein is digested and absorbed already in the bulbus part of the intestine.[8]Le et al. 2019. The study provided new basic insight into the anatomy, digestive, and immune function of the ballan wrasse’s intestine, and demonstrated that intestinal function and health are significantly affected by the composition of the feed.
Two ongoing projects, 10 and 11, Table 8, address brood stock nutrition and larval nutrition for farmed lumpfish and Ballan wrasse. Proper brood stock nutrition is essential for brood stock to develop good quality eggs, and proper nutrition in the early stages is essential for developing robust larvae. The goals for these projects are to contribute to better welfare and performance for farmed cleaner fish.
Wild-caught wrasse have a slightly different starting point than farmed ballan wrasse when it comes to feed and feeding strategies. Wild-caught wrasse are used to a varied food supply, and the different species have somewhat different diets. Farmed ballan wrasses are used to feed pellets and have fixed feeding times. The importance of feeding cleaner fish when they are being used in salmon cages is demonstrated in a paper by Skiftesvik et al. 2013. A lot of different feed and feeding strategies have been tried by farmers. Some work well for some of the species, but not for all. It will be favourable if all wrasses, farmed and wild caught, could be fed the same feed, and of course, the feed must satisfy the nutritional needs of the fish. One goal in the on-going project 31, Table 8, is to develop an optimized approach to feeding Ballan wrasse and lumpfish when they are in sea cages.
Feed for lumpfish
Supplementary feeding of cleaner fish deployed within commercial salmon pens is necessary to maintain the nutritional condition, welfare, and efficacy of the biological controls over the Atlantic salmon grow-out cycle, typically lasting 18–22 months. Therefore, a feed source adapted to the species feeding habits and to the salmon net-pens rearing environment has to be developed. It is important that lumpfish populations maintained in commercial salmon cages have access to a regular food source particularly in wintertime when naturally occurring food sources become scarce. This food source is vital to maintain healthy and robust populations. High or rapid growth can increase the risk of cataracts in salmon.[9]Ersdal et al. 2001. Previous studies on lumpfish[10]Imsland et al. 2018, Jonassen et al. 2017. have also found that high growth rate increased risk of developing cataracts. A recent study showed that fish fed with pellets had significantly higher growth compared with fish fed with feed blocks and that these fish had a high incidence of cataracts.[11]Imsland et al. 2019a. It is known that growth rates of small lumpfish are generally high, therefore one cannot rule out the possibility that high growth rates observed in lumpfish populations may contribute to the development of cataracts.
Rapid growth is not an aim for lumpfish used as cleaner fish. Imsland et al. (2016a) found that small lumpfish (initial size approx. 20 g) have a higher overall preference for natural food items, including sea lice, compared to larger fish. This makes slow to moderate and uniform growth of lumpfish more desirable than fast growth.[12]Imsland et al. 2016c. Additionally, lower feed input may encourage not fully satiated lumpfish to seek out alternative food sources such as attached sea lice due to their opportunistic feeding behaviour previously reported by Imsland et al. (2014a-c, 2015a).
Once lumpfish attain a mean weight over 200g their sea lice grazing behaviour generally decreases. If growth can be controlled the operational window of lumpfish will be extended. For example, data from a recent study (figure 9) showed that one group of lumpfish fed with feed blocks attained a mean weight of 53.8g at day 93 whilst the other group fed with commercial lumpfish pelleted feed attained a mean weight of 102.5g during the same period. This represents a difference of 48% lower mean weight for the fish fed with feed blocks. Using growth modelling for each group, the fish with pellets would have attained 200g at day 144 whilst for fish fed with feed blocks, 200g would have been attained at day 182. This represents a difference of 38 days extra when these fish are at their optimal size to graze sea lice. Controlling growth rates of lumpfish during the latter part of the hatchery phase and in commercial sea cages may allow for the prolongation of sea lice grazing behaviour. These growth differences allow for end users of lumpfish in commercial cages to alter their stocking strategies and potentially reduce the number of times restocking of lumpfish occurs, as well as enhancing sea lice grazing potential. These growth differences were also seen in the study of Imsland et al. (2020) further pinpointing this as a possible avenue to control growth of lumpfish in sea pens.
Footnotes
- ^ LT Barrett, pers. com.
- ^ barentswatch.no.
- ^ Imsland et al. 2018.
- ^ Espmark et al. 2020.
- ^ Ingólfsson and Kristjánsson 2002.
- ^ Imsland et al. 2015b, 2018b.
- ^ project 19, Table 8.
- ^ Le et al. 2019.
- ^ Ersdal et al. 2001.
- ^ Imsland et al. 2018, Jonassen et al. 2017.
- ^ Imsland et al. 2019a.
- ^ Imsland et al. 2016c.
Figure 9. Growth of lumpfish fed either feed blocks or commercial pelleted feed. The blue data points for both data series indicate modelled growth after day 93.
Recent studies at Gildeskål research station (Gifas) close to Bodø (Norway) suggest that feeding lumpfish with feed blocks may alleviate health issues due to enhanced nutritional intake and better controlled growth compared to fish fed with pelleted feed. The welfare of cleaner fish in cages is a prime concern and the focus of some of the fish welfare schemes. Lumpfish can lose condition within six weeks of transfer to sea cages. This can be alleviated by the supply of robust fish, and by providing a suitable supplementary feed source more suited to the species being fed. Using operational welfare indicators (OWIs) developed for this species, a recent study showed that lumpfish fed with feed blocks compared to fish fed with pelleted feed had consistently lower average scores indicating better health condition.[1]Figure 10, Imsland et al. 2020.
There is an urgent need for development of industry wide feeding strategies which facilitate the maintenance of good health status and controlled growth in lumpfish.
Footnotes
- ^ Figure 10, Imsland et al. 2020.
Figure 10. Mean health scores for lumpfish fed with feed blocks or commercial pelleted feed.
Recovery of cleaner fish from salmon cages
Cleaner fish are only used for one production cycle. The principle of fallowing to separate production cycles of fish is critical in terms of preventing transmission of disease. If the fish farmers have better success with good welfare and higher survival of cleaner fish, there will be a lot of cleaner fish left at the end of the salmon production cycle, and some projects have looked into how to use and re-use wrasse and lumpfish. An ongoing project[1]project 33, Table 8. aims to develop protocols for efficiently fishing out cleaner fish in connection with operations and slaughter of salmon, and to determine criteria for humane killing of cleaner fish. This can provide a basis for re-use of the fish, either for human consumption or through the extraction of valuable individual components. In a pilot project,[2]project 34, Table 8. controlled behaviour-based studies of light and colour preferences in lumpfish was carried out to develop passive recapture methods for use on a commercial scale. Laboratory-scale experiments demonstrated a high recapture rate using blue light as an attractant, but under commercial conditions the same method failed.
The main activity of the project was to develop a slaughter procedure that was suitable for industrial slaughter of wrasse and lumpfish that satisfies the requirement of animal welfare and at the same time safeguards the quality of the product as a possible food source. The project documented the sensitivity to electric current in all relevant species and identified the parameters (voltage / duration of exposure) that are necessary to stun and kill cleaner fish in accordance with regulations and concluded that existing commercial salmon anaesthetics were not suitable for anaesthesia and killing of cleaner fish.
The main aim of another ongoing project[3]project 35, Table 8. is to provide the knowledge necessary to turn farmed lumpfish into food for human consumption after they have completed their lice eating period. The project will “attain knowledge, address and solve challenges related to market, animal welfare through collection and sorting, nutritional content, processing methods, shelf-life, total biomass utilization, logistics, regulatory aspects and profitability through the whole value chain. The value-chain approach is important as the different aspects of innovation depend on information and solutions from other levels in the value chain. A secondary objective is to reveal solutions for downgraded raw material not suited for food, e.g. processing of high value products using bioprocessing methods, e.g. producing marine collagen powder.”
Table 8. Cleaner fish projects financed by FHF (Norwegian Seafood Research Fond), and NFR (The Research Council of Norway), not complete list. The link to FHF projects gives all project information about the projects, and reports and other outcomes from these projects will be there, or will be found there, as the projects progresses.
| No | Financed by | Project name | Period | Link |
| 1 | FHF prosjekt 901253 | Arts- og størrelsesseleksjon i leppefiskredskap | 2015-2016 | https://www.fhf.no/prosjekter/prosjektbasen/901253/ |
| 2 | FHF prosjekt 900554 | Produksjon av berggylt (LeppeProd) | 2010-2014 | https://www.fhf.no/prosjekter/prosjektbasen/900554/ |
| 3 | FHF prosjekt 900997 | Berggylt stamfiskhold | 2014-2015 | https://www.fhf.no/prosjekter/prosjektbasen/900997/ |
| 4 | FHF prosjekt 901135 | Kartlegging av berggyltens genom | 2015-2016 | https://www.fhf.no/prosjekter/prosjektbasen/901135/ |
| 5 | NFR HAVBRUK2 | Intestinal function and health in Ballan wrasse | 2015-2019 | https://prosjektbanken.forskningsradet.no/#/project/NFR/244170 |
| 6 | NFR HAVBRUK | Utvikling av oppdrett- og bruk av berggylte i lakseoppdrett til kontinuerlig kontroll med lakselus | 2007-2009 | https://prosjektbanken.forskningsradet.no/#/project/NFR/180028 |
| 7 | NFR HAVBRUK | Oppdrett av berggylte (Labrus bergylta) | 2003-2008 | https://prosjektbanken.forskningsradet.no/#/project/nfr/153261 |
| 8 | NFR HAVBRUK2 | Kontroll av kjønnsmodning hos rognkjeks | 2017-2021 | https://prosjektbanken.forskningsradet.no/#/project/NFR/269043 |
| 9 | FHF prosjekt 901418 | Reproduksjonsbiologi hos rognkjeks: En nøkkel til et vellykket avlsprogram (CYCLOBREED) | 2017-2021 | https://www.fhf.no/prosjekter/prosjektbasen/901418/ |
| 10 | FHF prosjekt 901561 | Optimalisert startfôring av rensefisk (STARTRENS) | 2019- 2021 | https://www.fhf.no/prosjekter/prosjektbasen/901561/ |
| 11 | FHF prosjekt 901562 | Kvalitetskriterier for rensefisk og effekten av stamfiskernæring (CleanLifeCycle) | 2019-2021 | https://www.fhf.no/prosjekter/prosjektbasen/901562/ |
| 12 | FHF prosjekt 901264 | Forsøk med dypp- og stikkvaksinering av rognkjeks i smittecelle og feltforsøk med oppfølging av vaksinert fisk i sjø | 2016-2017 | https://www.fhf.no/prosjekter/prosjektbasen/901264/ |
| 13 | NFR Nærings-PHD | Loss of salmon and cleaner fish, mapping of bacterial infections and preparatory steps for vaccine development | 2016-2019 | https://prosjektbanken.forskningsradet.no/#/project/NFR/260204 |
| 14 | NFR HAVBRUK2 | Basic immunology studies and development of tools to monitor immune responses in cleaner fish | 2015-2018 | https://prosjektbanken.forskningsradet.no/#/project/NFR/244396 |
| 15 | FHF prosjekt 901136 | Velferd hos rensefisk – operative indikatorer (RENSVEL) | 2015-2019 | https://www.fhf.no/prosjekter/prosjektbasen/901136/ |
| 16 | FHF prosjekt 901426 | Toleranse for transportstress og miljøoverganger hos berggylt og rognkjeks | 2017-2019 | https://www.fhf.no/prosjekter/prosjektbasen/901426/ |
| 17 | FHF prosjekt 901158 | Utvikling av transport- og mottaksprosedyrer for rognkjeks basert på kartlegging av miljø og stress | 2015-2017 | https://www.fhf.no/prosjekter/prosjektbasen/901158/ |
| 18 | FHF prosjekt 900979 | Bruk av rognkjeks i merd | 2014-2017 | https://www.fhf.no/prosjekter/prosjektbasen/900979/ |
| 19 | FHF prosjekt 900978 | Adferd og artssamspill i laksemerder | 2014-2018 | https://www.fhf.no/prosjekter/prosjektbasen/900978/ |
| 20 | NFR Nærings-PHD | Tilvenning og læringsevne hos rognkjeks i interaksjon med oppdrettslaks, med fokus på atferd og fysiologi | 2018-2022 | https://prosjektbanken.forskningsradet.no/#/project/NFR/283204 |
| 21 | NFR HAVBRUK2 | Optimal bruk av rognkjeks til avlusing av oppdrettslaks | 2016-2020 | https://prosjektbanken.forskningsradet.no/#/project/NFR/256199 |
| 22 | FHF prosjekt 900818 | Tapsårsaker og forbyggende tiltak | 2012-2014 | https://www.fhf.no/prosjekter/prosjektbasen/900818/ |
| 23 | FHF prosjekt 901188 | Utredning av dødelighet i forbindelse med akutt dødelighet/forhøyet dødelighet hos rognkjeks i 2015 | 2015-2016 | https://www.fhf.no/prosjekter/prosjektbasen/901188/ |
| 24 | FHF prosjekt 901120 | Analyse av sykdomsrelatert risiko forbundet med bruk av villfanget og oppdrettet rensefisk for kontroll av lakselus | 2015-2016 | https://www.fhf.no/prosjekter/prosjektbasen/901120/ |
| 25 | FHF prosjekt 901152 | Katarakt hos rognkjeks | 2015-2016 | https://www.fhf.no/prosjekter/prosjektbasen/901152/ |
| 26 | NFR Nærings-PHD | Identifisering og karakterisering av bakterielle sykdommer hos rensefisk (berggylt og rognkjeks) til bruk i biologisk kontroll av lakselus | 2012-2015 | https://prosjektbanken.forskningsradet.no/#/project/NFR/226695 |
| 27 | FHF prosjekt 901146 | Kunstig lys og rensefisk | 2015-2017 | https://www.fhf.no/prosjekter/prosjektbasen/901146/ |
| 28 | FHF prosjekt 901455 | Fullskala uttesting av Strømmen-rør for å dokumentere fiskevelferd og førebyggande effekt mot lakselus (Dette er prosjekt på laks, men man fikk høyere overlevelse på rensefisk i merder med Strømmen-rør). | 2018-2020 | https://www.fhf.no/prosjekter/prosjektbasen/901455/ |
| 29 | FHF prosjekt 901563 | Miljø og fôring for optimal helse og overlevelse av rensefisk i merd (OptiRens) | 2019-2021 | https://www.fhf.no/prosjekter/prosjektbasen/901563/ |
| 30 | FHF prosjekt 901652 | Effekt og velferd ved bruk av rensefisk og luseskjørt (EFFEKTIV) | 2020-2022 | https://www.fhf.no/prosjekter/prosjektbasen/901652/ |
| 31 | FHF prosjekt 901331 | Ernæringsbehov og fôring for optimal helse og overlevelse av rensefisk | 2017-2021 | https://www.fhf.no/prosjekter/prosjektbasen/901331/ |
| 32 | FHF prosjekt 901626 | Kan manglende tilgang på fòr eller indusert sult forårsake vektøkning hos rognkjeks (Cyclopterus lumpus) yngel? | 2020-2021 | https://www.fhf.no/prosjekter/prosjektbasen/ |
| 33 | FHF prosjekt 901560 | Gjenfangst, bedøvelse, avliving og etterbruk av rensefisk (CLEANCATCH) | 2019-2021 | https://www.fhf.no/prosjekter/prosjektbasen/901560/ |
| 34 | FHF prosjekt 901235 | Innfangning, avlivning og tilrettelegging for etterbruk av rensefisk – fra problem til ressurs: Forprosjekt | 2016-2017 | https://www.fhf.no/prosjekter/prosjektbasen/901235 |
| 35 | NFR HAVBRUK2 | From waste to food - sustainable exploitation of farmed lumpfish | 2020-2023 | https://prosjektbanken.forskningsradet.no/#/project/NFR/301494 |
| 36 | FHF prosjekt 901258 | Oppdatering av rensefiskveiledere | 2016-2017 | https://www.fhf.no/prosjekter/prosjektbasen/901258/ |
| 37 | FHF prosjekt 901647 | Effektiv og forsvarlig bruk av rensefisk: En kampanje for beste praksis bruk av rensefisk | 2020-2021 | https://www.fhf.no/prosjekter/prosjektbasen/901647/ |
| 38 | NFR HAVBRUK2 | The cleanerfish lumpfish (Cyclopterus lumpus L.)- Immunity, diseases and health | 2015-2019 | https://prosjektbanken.forskningsradet.no/#/project/NFR/244148 |
| 39 | FHF prosjekt 901692 | Årsak til dødelighet og tap av rensefisk (DOKUMENTAR) | 2021-2022 | https://www.fhf.no/prosjekter/prosjektbasen/901692/ |
| 40 | FHF prosjekt 901693 | Fôringsstrategiens påvirkning på ernæring og lusebeite-effektivitet hos rognkjeks i sjø (STRATEGI) | 2021-2023 | https://www.fhf.no/prosjekter/prosjektbasen/901693/ |
| 41 | FHF prosjekt 901694 | Optimal fôring av berggylt i laksemerd (OPTIfôr) | 2021-2023 | https://www.fhf.no/prosjekter/prosjektbasen/901694/ |
5. Health and welfare of cleaner fish in the net pens
All farming of animals in Norway must be done within the frames defined by the Norwegian Animal Welfare Act. In the beginning the welfare of cleaner fish was rarely discussed. This has changed, and there is an increased concern on the fate of the cleaner fish (see e.g. Rieber Mohn et al. 2021). Even though accurate data for the entire industry is lacking, data from The Cleaner fish campaign (see section 5.2) show that the mortality rate of cleaner fish is high. In many cases most of the cleaner fish die or disappear during the salmonid production cycle.
5.1 Regulatory framework
The Norwegian Ministry of Trade, Industry and Fisheries updated the regulations concerning transport, use and production of cleaner fish in 2018. The use of cleaner fish is regulated by several laws and regulations that include all aspects from breeding / fishing, transportation, and the actual usage of the fish in net pens. The regulations also have the purpose of securing good fish health and welfare.
- The Norwegian aquaculture Act.
Akvakulturloven - The Norwegian Animal Welfare Act
Dyrevelferdsloven - Regulations relating to Operation of Aquaculture establishments Akvakulturdriftsforskrifte
Akvakulturdriftsforskriften - Regulations of permission to farm fish.
Laksetildelingsforskriften - Regulations concerning abattoirs and processing plants for aquaculture animals.
Etableringsforskriften - Regulation concerning Transportation of Aquaculture Animals
Transportforskriften - Regulations concerning the capture and transportation of wild-caught fish to fish farms.
Forskrift om krav til fartøy - Regulations concerning the fishery of cleaner fish.
Forskrift om utøvelse av fiske - Regulations on internal control to meet the requirements of aquaculture legislation
IK-Akva - Regulations regarding sea lice on fish in fish farms.
Lakselusforskriften
5.2 Animal Welfare - The cleaner fish campaign
The Animal Welfare Act applies for farmed fish, including cleaner fish, based on the same principles as for any farmed terrestrial animal. It covers the entire chain of interaction with humans, from catching/fishing, transport, farming, use and killing.
The intention of the Animal welfare act is to promote good animal welfare and respect for animals. The act states that “Animals have an intrinsic value which is irrespective of the usable value they may have for man. Animals shall be treated well and be protected from danger of unnecessary stress and strains.”
Due to many reports of high mortality and poor animal welfare, The Norwegian Food Safety Authorities decided to do a campaign with coordinated controls. The aim of the campaign was to check if the industry was operating within the frames of the regulations and gain more knowledge on how good the welfare of the cleaner fish was. The campaign was called The cleaner fish campaign (Tilsynskampanjen med velferd for rensefisk), and was carried out in 2018 – 2019.
In addition to the use of cleaner fish in the sea cages, the campaign also included farming and fisheries. Inspections were carried out, and a total of 291 farms completed a questionnaire on their use of cleaner fish.[1]Stien et al. 2020, Størkersen and Amundsen 2019, NFSA 2020. Hatcheries reported an average loss from first count to delivery of almost 32 % for lumpfish and 66% for wrasses (i.e., Ballan wrasse).[2]Amundsen and Størkersen 2019. Once deployed in sea cages the farmers reported a registered mortality for lumpfish of 46 % and 37- 44% depending on wrasse species.[3]Stien et al. 2020, Størkersen and Amundsen 2019. However, the figures for mortality are not accurate. This is due to the fact that many dead lumpfish get caught in the net wall and never end up in the dead-fish collector at the bottom of the cage.[4]Geitung et al. 2020. On the other hand, some farmers may try to compensate for this by registering higher mortality numbers than they actually count. When asked how much they believe the mortality is for cleaner fish in sea cages with salmon, the farmers themselves estimated an average mortality of about 60 % for all species.[5]Stien et al. 2020, Størkersen and Amundsen 2019, NFSA 2020.
In the questionnaire, the farmers reported what they considered the most important cause of death of the cleaner fish. For wild-caught and farmed wrasses, de-lousing using non-medical methods was ranked as the primary cause of death (although cleaner fish should be removed before treatments, according to the regulations). Diseases were ranked second. For lumpfish, diseases were considered the most important factor, de-lousing the second. Thereafter, handling of fish and poor quality of the cleaner fish upon delivery was considered common causes of death.[6]Stien et al. 2020, Størkersen and Amundsen 2019, NFSA 2020.
The cleaner fish campaign has contributed to a better knowledge on the mortality rate of cleaner fish. The report and questionnaire can be accessed here: https://www.mattilsynet.no/fisk_og_akvakultur/akvakultur/rensefisk/rensefiskkampanje_20182019.30882
Having gained more knowledge, the NFSA can now focus on the most critical factors and encourage the industry to improve their practice.
The Animal Welfare Act states that: “Animals can only be kept if they can adapt to the method of keeping in a satisfactory way with regard to animal welfare.” A basic precondition for concluding that an animal species has adapted to the method of keeping must be that most of the animals survive the method of keeping. The high mortality rate that was shown in the cleaner fish campaign indicates that they are not able to adapt sufficiently to the environment in the farms. The NFSA has therefore announced that the fish farming industry must achieve a significant improvement in survival and welfare of the cleaner fish.
The Animal welfare act states that: “The animal keeper shall ensure that the animal receives good supervision and care, including securing that animals are protected from injury, disease, parasites and other dangers”. According to the cleaner fish campaign, delousing and diseases seems to be the main reasons of mortality. Special focus on these areas must therefore be considered as important.
The Aquaculture operation regulations requires that cleaner fish should be removed whenever salmon in sea cages are handled or treated, as this is an important factor causing mortalities in cleaner fish. The campaign confirmed that this is not always done. Additionally, the methods used to collect the cleaner fish from the net pens may not be efficient enough. To reduce the mortality, it is therefore crucial to improve methods and routines for welfare friendly collection of cleaner fish from the net pens.
The Aquaculture operation regulations, § 28 states that; “Fish should not be released into an aquaculture facility with an ongoing clinical disease outbreak if there is reason to believe that the released fish will become ill and be subjected to significant unnecessary discomfort.” However re-filling of cleaner fish into sea cages to replace the ones that are lost or have died, may happen even if there is diseased fish in the cage. To reduce the mortality caused by diseases it is important to obtain good disease detection routines to avoid adding cleaner fish to net pens with ongoing disease outbreak.
According to the Norwegian Animal Welfare Act, animals shall be treated well and be protected from danger of unnecessary stress and strains. It means that the placement of animals in an unnatural environment that may cause stress must be proven “necessary”. Keeping cleaner fish in captivity thus requires documentation of the effect. Although studied indicate that cleaner fish indicate effectively lower infestations at isolated farms, an overall a national scale study question their overall effect.[7]Barret et al. 2020.
Based on the results from the Cleaner Fish Campaign the NFSA has therefore stated that the use of cleaner fish cannot continue unless there is a significant improvement in welfare as well as a better documentation of cleaner fish as a de-lousing method.
Building on the results from the Cleaner Fish Campaign, the Norwegian Council for Animal Ethics came to a similar, but slightly more restrictive conclusion, in a recent recommendation (Norwegian Council for Animal Ethics, 2020) to the authorities: “The Norwegian Council for Animal Ethics means that todays practice, where millions of cleaner fish are used every year, is not justifiable, neither from an ethically nor an animal welfare point of view”. The council points at a lack of knowledge, both with regards to actual mortality, the effect of cleaner fish to remove lice and best practice to secure the cleaner fish species requirements for environment and handling conditions. They recommend limitations in the use of cleaner fish until better welfare and effect can be documented.
The need for improvement in the welfare of cleaner fish has also been laid down in the new (2021) Norwegian, governmental strategy for the aquaculture strategy,[8]Ministry of Trade, Industry and Fisheries, 2021. which states that “ . . . If the industry in the coming years cannot document in a satisfactory way that the cleaner fish can live good lives in the net cages and contribute significantly to fight salmon louse, the government see it as a natural consequence that the use of cleaner fish must be terminated”(own translation).
Footnotes
- ^ Stien et al. 2020, Størkersen and Amundsen 2019, NFSA 2020.
- ^ Amundsen and Størkersen 2019.
- ^ Stien et al. 2020, Størkersen and Amundsen 2019.
- ^ Geitung et al. 2020.
- ^ Stien et al. 2020, Størkersen and Amundsen 2019, NFSA 2020.
- ^ Stien et al. 2020, Størkersen and Amundsen 2019, NFSA 2020.
- ^ Barret et al. 2020.
- ^ Ministry of Trade, Industry and Fisheries, 2021.
5.3 Earlier assessment of mortality and welfare of cleaner fish in salmon pens
In project 22, Table 8, a systematic survey of causes of mortality in cleaner fish was performed, including collection and characterization of disease-causing bacterial isolates from cleaner fish. Infection models were made or used in vaccine development in Ballan wrasse and lumpfish. Two overviews of disease-causing organisms that occur in the various cleaner fish species was produced by the project. From the project’s final report: “In a period of 6 months, 934,935 cleaner fish were released, and 310,043 (33%) dead fish were registered. Registered mortality was lowest in Ballan wrasse (18%). For the other species registered mortality was from 32 to 48% and was highest in lumpfish. The most important causes of mortality were wounds and fin rot (19%), bacterial infections (14%), mechanical damage (6%) and sexual maturation (3%). Fish without a specific diagnosis accounted for 56%. Some of the first batches of wrasse had elements of sexual maturation, among which increased mortality was observed immediately after release. The most serious episodes of mortality after this were due to acute outbreaks of bacterial infection in the cleaner fish after release into cages (Pasteurella spp., Atypical Aeromonas salmonicida, Vibrio anguillarum) or operational problems in the facility related to weather, frequent uptake of dead salmon, net washing or drug treatment of salmon against salmon lice.”
Lumpfish aquaculture is faced with knowledge-gaps that result in poor health and welfare of the fish.[1]Imsland et al., 2019a-b; Powell et al., 2018. In a survey in five Atlantic salmon net pen sites with 79,000 lumpfish, conducted in 2013 in Norway an average of 48% mortality was reported, varying from 39% to 100%. Of the 13,864 dead lumpfish categorized regarding cause of death in the same survey from 2013 in Norway, 75% were categorized as bacterial infections.[2]Nilsen et al. 2014. The levels of mortality recorded vary widely between commercial sites and management strategies vary between users (see Figure 12). This may explain some of the differences observed.
Some of the loss may also be due to escapees. Wrasses are clever to find any holes in net pens and many can escape if only one thread is broken. It is also very important that right net mesh size is chosen to avoid cleaner fish to escape through the net. Goldsinny has the highest potential to escape, because they are small. In an experiment, 50 % escaped through the mesh used by the farmer for this batch of fish.[3]Woll et al. 2013. In a dedicated study, specifically looking at cleaner fish mortality in salmon sea cages, Geitung et al. (2020) found that while registered accumulated mortality after three months was 7 % for Ballan wrasse and 10% for Lumpfish, the actual loss calculated based on how many individuals that were left in the cages were almost 3 times as high for lumpfish and almost 8 times as high for Ballan wrasse. This discrepancy can be both due to escapees and that dead Ballan wrasse and lumpfish have been caught in the net wall and thereby never arrived into the dead fish collector in the bottom of the cage.
A recent study on skeletal deformities in cleaner fish found that 23% of the wild caught lumpfish had five or more deformed vertebra vs. only 1% in the wild caught ballan wrasse.[4]Fjelldal et al. 2020. Possible reasons for lumpfish ability to survive with severe deformities in the wild are that they feed on large slow-moving organisms and have a relative limited number of natural predators compared to wrasses of the same size. In contrast, the same study found that while none of the sampled cultured lumpfish had five or more deformed vertebrae, 17-53% of the sampled cultured ballan wrasse had severe spinal deformity. Fish with severe spinal deformity can have difficulties swimming properly and are therefore likely to experience low welfare if transferred to salmon sea cages. It should here be mentioned that also healthy lumpfish and ballan wrasse are poor swimmers compared to Atlantic salmon and therefore not recommended deployed at sites with high currents.[5]Hvas et al. 2021.
Animals have different natural lifespans, which needs to be considered when evaluating mortality of cleaner fish in salmon sea cages. In a large screening of the natural age composition of Skagerrak Corkwing (n=709), the average age was 1.4 years, with only one individual had reached four years.[6]Halvorsen et al. 2017. In Sweden, the maximum reported age is three years.[7]Uglem et al. 2000. In the light of this, the captive mortality of corkwing translocated from Skagerrak should be expected to be very high and the majority of corkwing would also have died within a years’ time in nature. On the other hand, the Cleaner fish campaign revealed that nearly 20% of the corkwing typically died within the first month after transfer to salmon sea cages.[8]Stien et al. 2020. The causes of natural mortality are not known, but includes disease, predation, but could also simply indicate a naturally low longevity – which applies both to captivity and nature. The absence of any old individual in these studies suggest that corkwing has a natural short lifespan in Skagerrak. However, corkwing in western Norway is considerably genetically differentiated from the short-lived populations in Skagerrak and can in rare cases become 8 years old. Goldsinny and Ballan have lower natural mortality and may attain more than 15 years of age in Norwegian waters,[9]Halvorsen et al. 2017. and all else being equal, should have lower mortality in captivity as well. It is therefore very important to record the species and the source of cleaner fish deployed – and we recommend that all farms should identify the species, sex and size of dead or diseased cleaner fish, information that will be of high value for improved welfare practices.
5.4 Operational Welfare Indicators (OWIs) and the development of better sampling regimes
Continuous health and welfare monitoring are essential to help identify when and what procedures and operations are affecting health and thus, allow us to adapt and improve practices.[1]Noble et al. 2019b, VKM. 2017. To ensure the adoption of health status monitoring in a standardized and more comparable way, practical and user-friendly approaches are necessary. Health scoring is becoming an ever-more common practice, and for this reason it is a valid method of inferring welfare status. Operational welfare indicators (OWI) have been developed to assess health and condition for lumpfish in recent studies.[2]Imsland et al. 2020, Rabadan et al. 2021. Both scoring systems focused on morphological health indicators. The assessment of health status is non-destructive, and lumpfish can easily be returned to their specific cage thus maintaining the population. The scoring systems used in these studies were shown to be robust and reliable and to give an accurate indication of the health and welfare status of the fish. Such scorings systems are proposed to be routinely used to screen the health and welfare of cleaner fish. Preferably via camera observations to avoid unnecessarily handling of the fish.
In project 15, Table 8, they found that plasma cortisol seemed to be a good indicator of stress for both lumpfish and Ballan wrasse, although the levels in lumpfish were much lower than in Ballan wrasse. In percentage terms, the increase in cortisol under stress was the same in lumpfish and Ballan wrasse. They also found that cleaner fish handle acute stress well, but fish welfare was negatively affected by long-term chronic stress, regardless of species. The results in the project and experience from experiments were, together with existing literature, summarized as a tool that breeders and others who assess cleaner fish health / welfare can benefit from. They produced fact sheets for Ballan wrasse and lumpfish. Both cover 34 OVIs (operational welfare indicators) and LABVIs (laboratory welfare indicators) for Ballan wrasse and lumpfish, respectively.
If OWIs indicate deterioration in health status and /or mortality rates are observed to increase, then detailed sampling regimes should be employed.[3]Kristiansen et al. 2020. Depending on the type of health issues reported, some sampling procedures can be better suited. These may be tissue samples for histological studies that can give more detailed information regarding health, gut condition, and even possible infection; blood sampling for haematology; plasma collection for analyses of stress biomarkers, immune status, and others; biopsies for PCR-analyses that can identify occurring infections the earliest.
5.5 Cataract status in lumpfish
Cataract is a significant problem in lumpfish. In a recent study, lumpfish fed with feed blocks had significantly less cataracts compared to the pelleted feed group.[1]Imsland et al. 2019b, Figure 11. The fish fed with feed blocks had 87% less cataracts after 93 days. The growth rates of small lumpfish are generally high and thus one cannot rule out the possibility that high growth rates observed in lumpfish populations may contribute to the development of cataracts. It may be that cataracts may be caused by not just high growth rates but also how efficiently the fish utilize the nutrients in the feed and particularly what type of nutrients the fish are consuming. Lumpfish with the highest prevalence may be consuming more salmon pelleted feed. Commercial salmon feed generally has less protein and higher fat content compared to feed blocks which are 100% marine profile with higher and lower inclusion levels of protein and fat respectively compared to salmon feed. In addition, salmon feed contains high inclusion levels of terrestrial ingredients. Using feed blocks results in less prevalence of cataracts and thus eye health is maintained. As lumpfish are visual feeders, maintaining eye health is of critical importance. There is an urgent need to identify potential triggers for cataract development in juvenile phases of production due to high prevalence of cataracts in commercial cages. A study on the relationship between cataracts and nutrition in lumpfish[2]Jonassen et al. 2017. suggests that cataracts in lumpfish may be related to primary or secondary disturbed nutrient metabolism or malnutrition which may cause osmotic imbalance and cataract development. Hence, a less nutrient-dense feed could be a better alternative for lumpfish, e.g. a moist feed.
Figure 11. Mean prevalence of cataracts of lumpfish fed with feed blocks (blue) or pelleted feed (yellow)
5.6 Infectious diseases of wild-caught wrasses introduced into the net pens
To avoid transmission of disease, it is as a general principle not possible to mix wild and farmed animals, within a farming unit, or even keep wild caught and farmed fish in proximity. Cleaner fish and salmonids may share pathogens.[1]Erkinharju et al. 2020, Table 9. Bacterial infections are the main cause of disease and mortality in cleaner fish and the number of recorded bacterial pathogens are expected to increase if the use of cleaner fish continues to grow. Pathogens may change virulence, and polyculture with a mixture of susceptible hosts and new pathogens may trigger host-switching and adaptation to new species and environment. In today’s farm situation / practice, the use of wild caught wrasse, with the repetitive long-distance transport and its unknown health status, may pose the highest risk of disease transfer. Today, salmonids are well protected through effective vaccines against many of these bacterial pathogens. However, when and if pathogens evolve and adapt to new species and environments, this could change. The disease status of wild-caught cleaner fish is, in general, still poorly known. A brief overview was presented by Institute of Marine Research, Risikorapporten 2017.[2]Mortensen et al. 2017. This report assessed the risk of disease transmission and spreading linked to translocations of cleaner fish. The report is updated annually.[3]Grefsrud et al. 2021a-b.
5.7 Viral diseases of wrasses
There is limited information on viral diseases of wrasses. Due to the extensive transportation of wild caught wrasses, unknown or undetected viruses may have been spread and introduced in naïve farming areas. There are some studies, focusing on viruses that are known to infect farmed fish, like infectious pancreatic necrosis virus (IPNV), viral haemorrhagic septicaemia virus (VHSV) and nerval necrosis virus (NNV):
The cleaner fish species do not seem to develop infectious pancreatic necrosis but may act as carriers of the virus.[1]Gibson and Sommerville 1996, Gibson et al. 2002. IPNV has a wide range of host species and may potentially be introduced and spread via cleaner fish.
Nerval necrosis virus (NNV) has been detected in wild Ballan, goldsinny and corkwing wrasse.[2]Korsnes et al. 2017. NNV has been described from several marine fish species and is known from disease outbreaks in farmed Atlantic cod and Atlantic halibut.[3]Patel et al. 2007; Grotmol et al. 1997. Different variants of the virus have been found in southern and northern parts of the distribution range of the wrasses. Transport of live wrasses will also represent a translocation of these viruses. NNV represents a hazard in farming of Ballan wrasse, potentially also lumpfish.
The IMR reports have particularly emphasized the risk of introducing viral haemorrhagic septicaemia virus (VHSV). It has also been recommended by VKM in 2019 to start screening cleaner fish for presence of VHSV (VKM 2019). VHSV adapts to new hosts and habitats. VHSV is a notifiable disease by international standard.[4]https://www.oie.int/en/what-we-do/animal-health-and-welfare/animal-diseases/ VHSV has been detected in wrasses in the Shetland Isles.[5]Munro et al. 2015. It is known that VHSV can transmit via food.[6]Ahne 1980, Schönherz et al. 2012. Small wrasses are sometimes eaten by salmon or rainbow trout, particularly during the period of starvation before slaughter. Salmonids may thus be exposed to VHSV through infected wrasse.
5.8 Bacterial pathogens of wrasses
Several pathogenic bacteria have been isolated from wrasse and lumpfish; strains of Vibrio anguillarum, Vibrio splendidus and atypical Aeromonas salmonicida are common (see e.g. Harkestad 2011, Johansen 2013, Treasurer 2012). An overview of mortality and causes was presented by Nilsen et al. (2014). Lumpfish and wrasse may suffer from pasteurellosis, caused by closely related Pasteurella-strains, but it is not clear whether transmission between the fish species occur.[1]Poppe et al. 2012, Alarcón et al. 2015. Tenacibaculum spp. and Moritella viscosa are frequently associated with skin ulcers in fish and have been isolated from both wild and farmed Ballan wrasse, and from lumpfish.
Footnotes
- ^ Poppe et al. 2012, Alarcón et al. 2015.
5.9 Parasites of wrasses
Amoebic gill disease (AGD) has caused severe problems in salmon farms in Norway since 2013. AGD is caused by the amoebae Paramoeba perurans. This amoeba has a marine reservoir, and has been isolated from several marine species, including Ballan wrasse and cuckoo wrasse.[1]VKM 2014. Infections and AGD has been observed in farmed Ballan wrasse[2]Karlsbakk et al. 2013. and lumpfish,[3]Bornø and Gulla 2016. and in corkwing, Ballan wrasse and lumpfish held as cleaner fish in net pens.[4]Nilsen et al. 2014, Karlsbakk 2015. Salmon can be infected with P. perurans isolated from Ballan wrasse,[5]Mo et al. 2014, Dahle et al. 2015 and Ballan wrasse can be infected with amoebae from salmon.[6]Dahle et al. 2015. Movements of fish with P. perurans represents a hazard of transmission to naïve fish.
5.10 Infectious diseases of lumpfish
When investigating mortality in connection with acute mortality / increased mortality in lumpfish in 2015,[1]project 23, Table 8. the results indicated that bacterial disease constitutes the most significant part of the problems related to mortality in lumpfish in Norway after release into the sea.
Bacterial infections are still considered to be the main cause for the high mortality rates observed in lumpfish.[2]Hjeltnes et al. 2017, Klakegg et al. 2020. Cataract represents a severe problem, as fish with severe cataracts have reduced ability to locate food resulting in reduced growth and weakened immunity and robustness of the fish which can result in high mortality rates (see above).
Project 24, Table 8 concluded that one can reduce the risk of disease and infection by only using farmed, vaccinated cleaner fish. Upon disease outbreak, traceability is very important to follow the same group of fish from production to farm stocking and thereby identify the probable time/place of infection. It is therefore necessary to introduce best practices relating to traceability of cleaner fish stocks and their provenance.
Lumpfish are affected by a series of diseases (Table 9). Often, more than one pathogen is isolated from diseased fish, making it difficult to verify the primary cause of disease and death. In addition, the cause of death may be multifactorial and influenced by poor nutritional input and/or inadequate feeding strategies. Extreme environmental conditions may also be a factor, and studies have shown that both low temperatures (< 4 oC,[3]Imsland et al. 2018.) and high temperatures (18 oC[4]Hvas et al. 2018.) can increase mortalities. High mortality can also occur after transfer to commercial cages due to poor handling and has been linked to chemical and mechanical de-lousing practices (e.g., Kousoulaki et al. 2018, Stien et al. 2020). Further, mortalities have been attributed to net cleaning, bath treatments or other operations.[5]VKM 2017, Marin Helse 2018, Stien et al. 2020.
Figure 12: Mean weekly percentage mortality rates of lumpfish from two commercial sites.
Table 9. Pathogens detected or associated with disease and mortalities in lumpfish.
| Type | Pathogen | Disease | Comments/reference |
| Bacteria | Vibrio anguillarum Vibrio spp. | Vibriosis | Vaccines available |
| Bacteria | Aeromonas salmonicida | Atypical furunculosis | Vaccine available |
| Bacteria | Moritella viscosa | Winter ulcer | Vaccine available |
| Bacteria | Pasteurella spp. | Pasteurellosis | Mortalities in hatcheries and sea cages |
| Bacteria | Pseudomonas anguilliseptica | Haemorrhagic septicaemia | Vaccine available |
| Bacteria | Tenacibaculum sp. | Ulcer | Vaccine available |
| Virus | Nodavirus | Viral nervous necrosis (VNN) | Unconfirmed observations |
| Virus | Flavivirus | Unknown, but associated with mortalities | Cyclopterus lumpus Virus (LFV/CLuV) Skoge et al. 2017 |
| Virus | Rhabdovirus | Viral Haemorrhagic Septicaemia (VHS) | Munro et al. 2015 Guðmundsdóttir et al. 2019 |
| Virus | Ranavirus | Unknown | Stagg et al. 2020 Sommerset et al. 2020 |
| Virus | Piscine Reovirus | Unknown | Reynolds et al. unpublished data |
| Parasite | Caligus elongatus | Ulcer | Øines et al. 2016 Sommerset et al. 2020 |
| Parasite | Paramoeba perurans | Amoebic gill disease | Sommerset et al. 2020 |
5.11 Bacterial diseases of lumpfish
Introduction of lumpfish as cleaner fish has led to an increasing number of observations of disease and poor health and welfare. Bacterial diseases are a main contributing cause to the high mortalities of lumpfish. The first pathogens causing disease in cultivated lumpfish were bacteria, and not surprisingly from the genus Vibrio. Vibriosis and atypical furunculosis were the first diagnosed diseases causing mass mortalities in lumpfish hatcheries and among lumpfish in commercial cages with salmonids. Another worrying emerging disease is pasteurellosis, caused by various strains of Pasteurella spp.[1]Ellul et al. 2019, Sandlund et al. 2021. Even though vaccines have been developed for most of the common bacterial pathogens, the number of bacteria causing disease challenges are steadily growing. Commercial vaccines are currently available as dip (larvae stage) or injection (juveniles) against Vibrios, Aeromonas, Pseudomonas, Moritella and Tenacibaculum, reducing the impact of these pathogens in cultivation of lumpfish.
Footnotes
- ^ Ellul et al. 2019, Sandlund et al. 2021.
5.12 Viral diseases of lumpfish
There are no reports of viruses causing mass mortality in cultivated lumpfish. However, several viruses which are known to cause challenges in marine fish species have been detected, pointing at a potential hazard. Rhabdovirus causing the notifiable disease viral haemorrhagic septicaemia (VHS) has been detected in lumpfish in Iceland.[1]Guðmundsdóttir et al. 2019. There has been a report on detection of Nodavirus in cultivated lumpfish (unpublished), which is a common marine virus found in several fish species. Similarly, piscine reovirus (PRV) has been detected in lumpfish kept in cohabitation with Atlantic salmon (unpublished data). Experimental challenge with infectious pancreatic virus (IPNV) has been demonstrated, but never detected in wild or cultivated lumpfish. Lumpfish flavivirus or Cyclopterus lumpus virus (LFV/CLuV) has been associated with inflammation and necrosis in liver,[2]Skoge et al. 2017. and gill pathology and anaemia has been reported in lumpfish with flavivirus.[3]Johansen et al. 2019. There are also some new virus types recently described, as Cyclopterus lumpus Coronavirus (CLuCV) and Cyclopterus lumpus Totivirus (CLuTV), although the significance of these viruses is unknown.[4]Sommerset et al. 2020. Ranavirus has been detected in lumpfish at the Faroe Islands and Iceland.[5]Stagg et al. 2020. These pathogens should therefore be monitored.
5.13 Lumpfish parasites
Lumpfish are farmed, and this may limit the hazard of introducing parasites commonly found in wild fish into the farming facilities. However, there are concerns regarding parasites in salmonid aquaculture and parasites horizontally transferred from wild fish populations. One parasite in focus is Caligus elongatus, which is a fish louse also infecting salmonids. There have been some reports of lumpfish with extensive C. elongatus infections in the northern part of Norway,[1]Sommerset et al. 2020. suggesting this parasite might cause an increasing challenge in lumpfish. In addition, this parasite also infects salmonids, suggesting a potential transmission of parasites between these two fish spices. However, C. elongatus from lumpfish seems to be host specific, suggesting a reduced risk of transfer.[2]Øines et al. 2006. Another parasite detected in lumpfish, which is causing mortalities in salmonids, is Paramoeba perurans, causing amoebic gill disease. This parasite has been detected in lumpfish kept in cages with infected salmon as well as tanks onshore.
5.14 Transfer of pathogens between cleaner fish and salmonids?
The risk of transferring pathogens from cleaner fish to the farmed salmonids has been assessed by The Norwegian Scientific Committee for Food and Environment (Vitenskapskomiteen format og miljø, VKM.[1]VKM 2017, 2019. The general finding was that cleaner fish have their own, specific pathogens. Some of these appear to be host species-specific, but others may be promiscuous regarding host species. Ten viruses were identified as possible hazards to farmed salmonids: Piscine myocarditis virus (PMCV), Viral haemorrhagic septicaemia virus (VHSV), Infectious salmon anaemia virus (ISAV), Salmonid alphavirus (SAV), Infectious pancreas necrosis virus (IPNV), Nodavirus, Piscine orthoreovirus (PRV), Lymphocystis virus, Lumpfish ranavirus, and Lumpfish flavivirus. Ten bacterial agents were identified as possible hazards: Aeromonas salmonicida subsp. salmonicida, atypical Aeromonas salmonicida, Vibrio anguillarum (primarily serotypes O1 and O2a), Vibrio ordalii, Vibrio spp., Pasteurella spp., Pseudomonas anguilliseptica, Piscirickettsia salmonis, Moritella viscosa, and Tenacibaculum spp. Farmed salmon are however vaccinated against V. anguillarum, Aliivibrio salmonicida, Moritella viscosa and typical A. salmonicida. Two parasites were identified as possible hazards: Paramoeba perurans and Caligus elongatus.
A recent challenge experiment conducted at the Institute of Marine Research, tested Atlantic salmon and lumpfish susceptibility to two isolates of Pasteurella spp. isolated from diseased Atlantic salmon and lumpfish, respectively. Only lumpfish developed clinical signs of disease, suggesting high susceptibility in lumpfish to Pasteurella-isolates originating from both Atlantic salmon and lumpfish.[2]Sandlund et al. 2021. A study conducted at Gifas in Nordland county in Norway, lumpfish kept in cohabitation with salmon were screened sequentially through a period for known pathogens. In this study, piscine reovirus (PRV) was detected in lumpfish only after transfer to cages with salmon, and with increasing load of virus throughout the experimental period. PRV was also detected in salmon at all sampling points, suggesting both fish species were infected by the virus.[3]Reynolds et al. unpublished data. This finding suggests a transfer of PRV had occurred at the marine growth site. These two studies may demonstrate the potential risk of pathogen transfer between lumpfish and salmonids.
5.15 Immunology and vaccine development
It became clear early on that farmed lumpfish were very prone to diseases when they were introduced into sea cages and vaccine development was initiated early.
In project 12, Table 8, they tried out different vaccination methods, but could not conclude because most of the fish they put out in the sea cages disappeared or died. In project 38, Table 8, they said the situation with large scale disease outbreaks among the lumpfish, caused by Pasteurella-like bacteria, Vibrio species and atypical Aeromonas salmonicida, clearly showed the need for immune prophylaxis like vaccination and/or immune stimulation. Their goals were to develop protocols for anaesthesia and antibacterial treatment.
In project 13, Table 8, the objective was to 1) source and map the prevalence and frequency of the predominant bacterial species on farms, including evaluations of various strategies such as custom or autogenous vaccination and treatments toward an improved natural bacterial biota and 2) to develop pilot vaccines for lumpfish and wrasse species and to test those vaccines in research aquarium facilities with candidate pathogenic bacterial species for vaccine development with the intention to improve the overall health and welfare of the cleaner fish.
Project 24, Table 8 identified the primary pathogens; atypical Aeromonas salmonicida, Vibrio splendidus and V. tapetis. Characterization of the pathogenic properties and the antigenic diversity within these groups were considered a necessity to form the basis for selecting candidates for inclusion in future vaccines for cleaner fish.
Project 14, Table 8, revealed key immune genes and characteristics of the immune system in wrasse and lumpfish by transcriptome and immunohistochemistry studies.[1]Bilal et al. 2016, 2018, 2019, Erkinharju et al. 2019.
Footnotes
- ^ Bilal et al. 2016, 2018, 2019, Erkinharju et al. 2019.
5.16 Suggested activities aiming at improving welfare and survival in captivity
To fulfil the requirements signalized by the authorities, it is necessary to:
- show that it is possible to give the different cleaner fish species a good life in the net cages.
- improve best practice manuals and guidelines for cleaner fish.
- identify the causal mechanisms for disease and mortality in cleaner fish.
- develop methodology for improved health and welfare in hatcheries, and strategies to improve their robustness when transferred to sea cages.
- develop methodologies and system that increase the welfare of cleaner fish in sea cages.
- Develop vaccines for the most prevalent and detrimental diseases
- obtain documentation of how efficient the different species are, and if the use of cleaner fish contributes significantly to reduce the number of salmon lice.
Some specific improvements to best practice and guidelines include:
- Optimal size of lumpfish at the time of transfer
- Best time for transfer to the net pens
- Optimal shelters and resting areas
- Technical solutions to separate salmonids and cleaner fish
- Optimized feed and feeding
To know whether farmers in the future can improve the welfare of the cleaner fish it is essential for the authorities, and the farmers, to have reliable numbers of mortality and survival of cleaner fish in the farms. It is thus urgent:
- to develop systems for more accurate registration of mortality and monitoring welfare of cleaner fish in salmon sea cages.
New systems for real time surveillance of mortality, for example based on machine vision and artificial intelligence, can contribute to better documentation of fish health and welfare.
Today’s reporting system of biomass and fish mortality to the government is also outdated, making the data difficult to analyse. There should therefore
- be developed new reporting systems between the farmers and the government.
Toda systems is cumbersome in that the farmers must simplify the data they have in their own computer systems to make it fit into the reporting standard. Machine to machine (automatic reporting from system to system) reporting may be one way to both reduce the farmers reporting burden and ensure more accurate data.
6. Epilogue
We have now entered the United Nations Decade of Ocean Science for Sustainable Development. Though this project, we have learnt that collaboration across national borders is essential for progress towards sustainable management of fishing and using cleaner fish, and that managers and legislators working on the various aspects should be aware of the wider implications of their management decisions. This can be the first step towards a holistic management of cleaner fish. We also hope that our review of the specific challenges for cleaner fish can be useful for a wider assessment and evaluation of the strategies for mitigating the sea-lice problem and improve sustainability of salmon aquaculture, in Norway and elsewhere.
As illustrated in the previous chapters, the use of cleaner fish comes with several complex challenges whose impacts extends from ecosystems to the welfare of individual fish. In addition, several challenges are intrinsically linked, and mitigation may involve difficult trade-offs for managers. For instance, sourcing cleaner fish from a larger geographical area may reduce the exploitation pressure on local populations but comes with a risk of introducing new diseases and genetic pollution. Policy makers do also have to consider the positive socio-economic effects of these fisheries on coastal communities, as small-scale fisheries have been in decline in most parts of Northern Europe. Regarding fish welfare, there are valid arguments to reduce or even prohibit the use of cleaner fish if not fish welfare improves, but this must be carefully considered, as it could induce an increase of mechanical or chemical methods which have different negative impacts on salmon welfare or other marine organisms affected by de-lousing agents. However, there is currently a lot of research effort with the goal to increase survival and welfare for the different cleanerfish species used. It has been beyond the scope and intention of this report to weigh the pros and cons of using cleaner fish to other alternatives. Rather, our approach has been solution-oriented, highlighting the key challenges and knowledge gaps for each challenge, exchanging experiences and providing specific advice for scientists, managers and farmers. We have done our best to identify the critical knowledge gaps that need to be filled in order to achieve an understanding of the risks and impacts of the current practice of fishing and using cleaner fish.
For ecosystem impacts, it is important to continue and expand rigorous assessment of the wild populations of wrasse through annual surveys, so that natural and fisheries induced changes in the populations can be detected early and allow managers to respond with the appropriate measures. A key consideration is how to assess and mitigate the risks associated with long-distance transportation of cleaner fish. There is evidence that this have caused hybridization between isolated wrasse populations, but the consequences for fitness and scope for adaptation in the affected populations is still unknown. Transmission of diseases and parasites, and introduction of new species through the transport water may cause severe, but highly uncertain impacts on ecosystems in recipient areas, as reviewed in recent risk assessments.[1]VKM 2019, Grefsrud et al. 2021. Fortunately, there is so far little evidence of disease transmission between farmed salmon in sea cages and wild fish, but if it happens, the consequences can be irreversible and have wide ramifications.
In the last decade, the demand for cleaner fish has been considerably higher than supply of both wild sourced and farmed wrasse and lumpfish. However, this could rapidly change through breakthrough in new technology for removing lice, but also due to stricter welfare regulations. Although cultured cleaner fish has by now surpassed wild-caught cleaner fish in numbers, the demand for wild-caught wrasse is likely to persist until a large-scale production of cultured Ballan wrasse has been established and can compete with the prices of wild-caught wrasses. It is unknown when, or if, this will happen, and we believe it should not be used as a justification for not to dedicate resources in improving assessments and management of wild wrasse populations.
Regarding the use of cleaner fish, there are considerable welfare challenges with need urgent attention. The future use of cleaner fish depends on a proactive and transparent industry that is dedicated to rapid improvement of the current situation for cleaner fish in captivity. Practical solutions or inventions to solve these challenges should be well documented, and scientifically evaluated, which require close collaboration between the relevant research institutes and managers. In the same way, there is a need to better document the effectiveness of the various cleaner fish species under different conditions. There are several ongoing research projects dedicated to these challenges, spurred by the recent focus on welfare and effectiveness of cleaner fish. We welcome the public debate on the ethical dilemmas and welfare challenges for cleaner fish, which we believe can be a catalyst for improved sustainability.
Footnotes
- ^ VKM 2019, Grefsrud et al. 2021.
7. References
Anon 2020. Sluttrapport: Nasjonalt tilsynskampanje 2018/2019: Velferd hos rensefisk. The Norwegian Food Authorities, Norway
Ahne W (1980). Experimental infection of pike (Esox lucius) with Egtvedvirus. Tierarztl Umsch 35: 225-229.
Alarcón M, Gulla S, Røsæg MV, Rønnseth A, Wergeland H, Poppe TT, Nilsen H, Colquhoun DJ (2015). Pasteurellosis in lumpsucker Cyclopterus lumpus, farmed in Norway. Journal of Fish Diseases 39(4): 489–495. https://doi.org/10.1111/jfd.12366
Almada F, Casas L, Francisco SM, Villegas-Ríos D, Saborido-Rey F, Irigoien X, Robalo JI (2016). On the absence of genetic differentiation between morphotypes of the ballan wrasse Labrus bergylta (Labridae). Marine Biology 163: 86. https://doi.org/10.1007/s00227-016-2860-8
Alvsvåg J (1993). Fødeval, vekst og energiallokering hos grasgylt (Centrolabrus exoletus L.) og grønngylt (Symphodus melops L.) (Pisces: Labridae) på vestkysten av Noreg, og diettoverlapping med O og l-gruppe torsk (Gadus morhua L.). (Master's thesis, University of Bergen). In Norwegian.
Alonzo, S. H., Mangel, M. (2004). The effects of size-selective fisheries on the stock dynamics of and sperm limitation in sex-changing fish. Fishery Bulletin, 102, 1–13. Amundsen VS, Størkersen KV (2019). Rensefiskoppdrett 2019 : Resultat av spørreundersøkelse til rensefiskoppdrettere. Studio Apertura ISBN/ISSN: 978-82-7570-587-5. In Norwegian.
Barrett LT, Oppedal F, Robinson N, Dempster (2020). Prevention not cure: a review of methods to avoid sea lice infestations in salmon aquaculture. Reviews in Aquaculture, 12(4): 2527–2543. https://doi.org/10.1111/raq.12456
Barrett LT, Overton K, Stien LH, Oppedal F, Dempster T (2020). Effect of cleaner fish on sea lice in Norwegian salmon aquaculture: a national scale data analysis. International Journal for Parasitology, 50(10-11): 787–796. https://doi.org/10.1016/j.ijpara.2019.12.005
Baskett, ML, Barnett, LAK. (2015). The Ecological and Evolutionary Consequences of Marine Reserves. Annual Review of Ecology, Evolution, and Systematics, 46: 49–73. https://doi.org/10.1146/annurev-ecolsys-112414-054424
Besnier F, Kent M, Skern-Mauritzen R, Lien S, Malde K, Edvardsen R B, … Glover KA (2014). Human-induced evolution caught in action: SNP-array reveals rapid amphi-atlantic spread of pesticide resistance in the salmon ecotoparasite Lepeophtheirus salmonis. BMC genomics, 15, 937.
Bjordal, A (1988). Cleaning symbiosis between wrasses (Labridae) and lice infested salmon (Salmo salar) in mariculture. International Council for the Exploration of the Sea CM1988, F:17. Available from: https://imr.brage.unit.no/imr-xmlui/handle/11250/104474
Bjordal, A (1990). Sea lice infestation on farmed salmon: possible use of cleaner-fish as an alternative method for de-lousing. Canadian Technical Report in Fisheries and Aquatic Sciences 1761: 85–89.
Bjordal, A (1992). Cleaning symbiosis as an alternative to chemical control of sea lice infestations of Atlantic salmon. In: Thorpe JE, Huntingford FA (Eds.). The importance of feeding behaviour for the efficient culture of salmonid fishes World Aquaculture Workshops, World Aquaculture Society, pp. 53–60.
Blanco Gonzalez E, Knutsen H, Jorde PE (2016). Habitat discontinuities separate genetically divergent populations of a rocky shore marine fish. PLoS ONE 11. https://doi.org/10.1371/journal.pone.0163052
Bollinger PJ (2020). Biological control of salmon lice: A critical analysis of knowledge production and development in the Norwegian cleaner fish industry. Master thesis, University of Life Sciences. https://nmbu.brage.unit.no/nmbu-xmlui/handle/11250/2683461
Bolstad GH, Hindar K, Robertsen G, Jonsson B, Sægrov H, Diserud OH, Fiske P, Jensen AJ, Urdal K, Næsje TF, Barlaup BT, Florø-Larsen B, Lo H, Niemelä E, Karlsson S (2017). Gene flow from domesticated escapes alters the life history of wild Atlantic salmon. Nature Ecology & Evolution 1(5): 0124. https://doi.org/10.1038/s41559-017-0124
Bornø G, Gulla S (2016). Helsesituasjonen hos rensefisk. S. 70-73 i (Hjeltnes m.fl. red). Fiskehelserapporten 2015. Oslo: Veterinærinstituttet.
Bourlat, S. J., Faust, E., Wennhage, H., Wikström, A., Rigby, K., Vigo, M., … André, C. (2021). Wrasse fishery on the Swedish West Coast: towards ecosystem-based management. ICES Journal of Marine Science, 78, 1386–1397.
Breen M (1996). Field observation of the cleaning of Ballan wrasse and torment feeding. In: Wrasse Biology and use in Aquaculture. (eds M.D.J. Sayer, M.J. Costello and J.W. Treasurer). Oxford: Fishing News Books, pp 55–59.
Burka JF, Hammell KL, Horsberg TE, Johnson GR, Rainnie DJ, Speare DJ (1997). Drugs in salmonid aquaculture - A review. Journal of Veterinary Pharmacological Therapy 20: 333–349. https://doi.org/10.1046/j.1365-2885.1997.00094.x
Burridge L, Weis JS, Cabello F, Pizarro J, Bostick K (2010). Chemical use in salmon aquaculture: A review of current practices and possible environmental effects. Aquaculture 306: 7–23. https://doi.org/10.1016/j.aquaculture.2010.05.020
Carl H, Møller PR (Eds.). Atlas over danske saltvandsfisk. Statens Naturhistoriske Museum. Online-udgivelse, december 2019. [https://fiskeatlas.ku.dk/]. In Danish.
Casas L, Saenz-Agudelo P, Villegas-Ríos D, Irigoien X, Saborido-Rey F (2021). Genomic landscape of geographically structured colour polymorphism in a temperate marine fish. Molecular Ecology 30: 1281–1296. https://doi.org/10.1111/mec.15805
Collins MAJ(1978). Experiments on the hatching period of the eggs of the lumpfish (Cyclopterus lumpus L. in Newfoundland waters. Naturaliste Canadien 105(3): 169–171.
Council for Animal Ethics (2020): Is it possible to use cleanerfish in an ethical and welfare-sound way in Norwegian fish farms? (In Norwegian: Er det mulig å bruke rensefisk på en etisk og velferdsmessig forsvarlig måte i norske oppdrettsanlegg?) https://www.radetfordyreetikk.no/er-det-mulig-a-bruke-rensefisk-pa-en-etisk-og-velferdsmessig-forsvarlig-mate-i-norske-oppdrettsanlegg/
Costello M (1991). A review of the biology of wrasse (Labridae) in Northern Europe. Progress in Underwater Science 16: 29–51.
Curtin S, Henly L, Stewart J E (2020). Three Year Comprehensive Review of the Live Wrasse Fishery in Devon and Severn IFCA’s District. Devon and Severn IFCA.
D’Arcy J, Mirimin L, FitzGerald R (2013). Phylogeographic structure of a protogynous hermaphrodite species, the ballan wrasse Labrus bergylta, in Ireland, Scotland, and Norway, using mitochondrial DNA sequence data. ICES Journal of Marine Science 70: 685–693. https://doi.org/10.1093/icesjms/fst018
Dahle OM, Nylund A, Karlsbakk E, Andersen L, Blindheim S, Glosvik H, Breck O (2015). Smitteoverføring av Paramoeba perurans mellom laks og berggylt - bruk av to klonale isolat fra henholdsvis laks og berggylt. Abstract, Frisk Fisk, Tromsø.
Darwall WRT, Costello MJ, Donnelly R, Lysaght S (1992). Implications of life-history strategies for a new wrasse fishery. Journal of Fish Biology 41: 111–123.
Davenport, J., Lønning, S., (1980). Oxygen uptake in developing eggs and larvae of the cod, Gadus morhua L. Journal of Fish Biology, 16: 249–256.
Davenport, J. (1985). Synopsis of biological data on the lumpsucker, Cyclopterus lumpus (Linnaeus, 1758). FAO Fisheries Synopsis, 147. 31 pp. Available from: http://www.fao.org/3/ap950e/ap950e.pdf
Davies S (2016) A review of wrasse ecology and fisheries interactions., Book September 2016. D&S IFCA paper.
Deady S, Fives JM (1995a). Diet of Ballan wrasse, Labrus bergylta, and some comparisons with the diet of corkwing wrasse, Crenilabrus melops. Journal of the Marine Biological Association of the United Kingdom 75: 651–665.
Deady S, Fives JM (1995b). The diet of corkwing wrasse, Crenilabrus melops, in Galway Bay, Ireland, and in Dinard, France. Journal of the Marine Biological Association of the United Kingdom 75: 635–649.
Deady S, Varian SJA, Fives JM (1995). The use of cleaner-fish to control sea lice on two Irish salmon (Salmo salar) farms with particular reference to wrasse behaviour in salmon cages. Aquaculture 131(1-2): 73–90.
Dipper FA, Pullin RSV (1979). Gonochorism and sex-inversion in British Labridae (Pisces). Journal of Zoology, London 187: 97–112.
Dipper FA, Bridges CR, Menz A (1977). Age, growth and feeding in the ballan wrasse Labrus bergylta Ascanius 1767. Journal of Fish Biology, 11 (2): 105–120.
DSIFCA. 2020. New Potting Permits Being Issued. https://www.devonandsevernifca.gov.uk/Latest-news/New-Potting-Permits-Being-Issued (Accessed 10th of August 2020).
Egidius E (1987). Import of furunculosis to Norway with Atlantic salmon smolts from Scotland. ICES CM Documents. 1987(F:8).
Eliasen K, Danielsen E, Johannesen Á, Joensen LL, Patursson EJ (2018). The cleaning efficacy of lumpfish (Cyclopterus lumpus L.) in Faroese salmon (Salmo salar L.) farming pens in relation to lumpfish size and seasonality. Aquaculture, 488: 61–65. https://doi.org/10.1016/j.aquaculture.2018.01.026
Eriksen E, Durif CMF, Prozorkevich D (2014). Lumpfish (Cyclopterus lumpus) in the Barents Sea: development of biomass and abundance indices, and spatial distribution. ICES Journal of Marine Science 71: 2398–2402. https://doi.org/10.1093/icesjms/fsu059
Eriksson BK, Sieben K, Eklf J, Ljunggren L, Olsson J, Casini M, Bergstrøm, U (2011). Effects of altered offshore food webs on coastal ecosystems emphasize the need for cross-ecosystem management. Ambio 40(7): 786–797. doi: 10.1007/s13280-011-0158-0
Erkinharju T, Dalmo RA, Hansen M, Seternes T (2020). Cleaner fish in aquaculture: review on diseases and vaccination. Reviews in Aquaculture. 1–49. https://doi.org/10.1111/raq.12470
Ersdal C, Midtlyng PJ, Jarp J (2001). An epidemiological study of cataracts in seawater farmed Atlantic salmon Salmo salar. Diseases of Aquatic Organisms 45: 229–236.
Fallang A, Ramsay JM, Sevatdal S, Burka JF, Jewess P, Hammell KL. Horsberg TE (2004). Evidence for occurrence of an organophosphate-resistant type of acetylcholinesterase in strains of sea lice (Lepeophtheirus salmonis Krøyer). Pest Management Science 60(12): 1163–1170. doi: 10.1002/ps.932.
Faust E, DeWit P, Wennhage H, Selander E, Strand Å, Halvorsen KT, Mattingdal M, Jorde PE, Blanco-Gonzalez E, Knutsen H, André C. Management of a harvested species with low genetic diversity and weak population structure: the corkwing wrasse. In preparation.
Faust E, Jansson E, André C, Halvorsen KT, Dahle G, Knutsen H, Quintela M, Glover KA. (2021). Not that clean: aquaculture mediated translocation of cleaner fish lead to hybridization and introgression on the northern edge of the species's range. Evolutionary Applications. https://doi.org/10.1111/eva.13220
Faust E, Halvorsen KT, Andersen P, Knutsen H, André C (2018). Cleaner fish escape salmon farms and hybridize with local wrasse populations. Royal Society Open Science 5: 171752. https://doi.org/10.1098/rsos.171752
Fjelldal PG, Madaro A, Hvas M, Stien LH, Oppedal F, Fraser TWK (2020). Skeletal deformities in wild and farmed cleaner fish species used in Atlantic salmon Salmo salar aquaculture. Journal of fish biology 98(4): 1049-1058. https://doi.org/10.1111/jfb.14337
Fjørtoft HB, Nilsen F, Besnier F, Espedal PG, Stene A, Tveten A-K, Bjørn PA, Aspehaug T, Glover KA (2020), Aquaculture-driven evolution: distribution of pyrethroid resistance in the salmon louse throughout the North Atlantic in the years 2000–2017, ICES Journal of Marine Science, Volume 77, Issue 5, September 2020, Pages 1806–1815, https://doi.org/10.1093/icesjms/fsaa062
Froese R (2004) Keep it simple: Three indicators to deal with overfishing. Fish and Fisheries 5: 86–91.
Funk WC, McKay JK, Hohenlohe PA, Allendorf FW (2012). Harnessing genomics for delineating conservation units. Trends in Ecology & Evolution 27(9): 489–496. doi.org/10.1016/j.tree.2012.05.012
Garcia-Mayoral E, Olsen M, Hedeholm R, Post S, Nielsen EE, Bekkevold D (2016). Genetic structure of West Greenland populations of lumpfish Cyclopterus lumpus. Journal of Fish Biology, 89(6): 2625–2642. doi.org/10.1111/jfb.13167
Geitung L, Oppedal F, Stein LH, Dempster T, Karlsbakk E, Nola V, Wright DW (2019). Snorkel sea-cage technology decreases salmon louse infestation by 75% in a full-cycle commercial test. International Journal of Parasitology 49 (11): 843-846. https://doi.org/10.1016/j.ijpara.2019.06.003
Geitung L, Wright DW, Oppedal F, Stien LH, Vågseth T, Madaro A (2020). Cleaner fish growth, welfare and survival in Atlantic salmon sea cages during an autumn-winter production. Aquaculture 528: 735623. https://doi.org/10.1016/j.aquaculture.2020.735623
Gibson DR, Sommerville C (1996). The potential for viral problems related to the use of wrasse in the farming of Atlantic Salmon. s 240-246 I: Sayer MDJ, Treasurer JW, Costello MJ (red.): Wrasse biology and use in aquaculture. Fishing news books. Blackwell, Oxford.
Gibson DR, Smail DA, Sommerville C (2002). Infectious pancreatic necrosis virus: experimental infection of goldsinny wrasse, Ctenolabrus rupestris L. (Labridae). Journal of Fish Diseases 21: 399–406.
Gjøsaeter, J. (2002). Fishery for goldsinny wrasse (Ctenolabrus rupestris) (Labridae) with pots along the Norwegian Skagerrak coast. Sarsia, 87, 83–90
Glover KA, Solberg MF, Mcginnity P, Hindar K, Verspoor E, Coulson MW, Hansen MH, Araki H, Skaala Ø, Svåsand T (2017). Half a century of genetic interaction between farmed and wild Atlantic salmon: Status of knowledge and unanswered questions. Fish and Fisheries 18:890–927. https://doi.org/10.1111/faf.12214
Glover KA, Wennevik V, Hindar K, Skaala Ø, Fiske P, Solberg MF, Diserud OH, Svåsand T, Karlsson S, Andersen LB, Grefsrud ES (2020). The future looks like the past: Introgression of domesticated Atlantic salmon escapees in a risk assessment framework. Fish and Fisheries 2020;00:1–15. doi.org/10.1111/faf.12478
Gonzalez EB, Knutsen H, Jorde PE (2016) Habitat discontinuities separate genetically divergent populations of a rocky shore marine fish. PloS one 11:e0163052. https://doi.org/10.1371/journal.pone.0163052
Goulet D, Green JM, Shears TH (1986). Courtship, spawning, and parental care behavior of the lumpfish, Cyclopterus Lumpus L., in Newfoundland. Canadian Journal of Zoology-Revue Canadienne De Zoologie 64(6): 1320-1325.
Goulet D, Green JM (1988). Reproductive success of the male lumpfish (Cyclopterus lumpus L.) (Pisces: Cyclopteridae): evidence against female mate choice. Canadian Journal of Zoology 66, 2513–2519
Gozlan RE, Peeler EJ, Longshaw M, St-Hilaire S, Feist SW (2006). Effect of microbial pathogens on the diversity of aquatic populations, notably in Europe. Microbes and Infection 8(5):1358–1364. doi.org/10.1016/j.micinf.2005.12.010
Gravestock V (2018) HRA - Studland to Portland SAC - Fish traps. Southern IFCA
Grefsrud, E.S., Karlsen, Ø., Kvamme, B.O., Glover, K., Husa, V., Hansen, P.K., Samuelsen, O.B., Sandlund, N., Stien, L.H. and Svåsand, T. 2021a. Risikorapport norsk fiskeoppdrett - risikovurdering (in Norwegian), Rapport fra Havforskningen, 2021-8, 198 pp. https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2021-8
Grefsrud, E.S., Karlsen, Ø., Kvamme, B.O., Glover, K., Husa, V., Hansen, P.K., Samuelsen, O.B., Sandlund, N., Stien, L.H. and Svåsand, T. 2021b. Risikorapport norsk fiskeoppdrett 2021 - kunnskapsstatus (in Norwegian), Rapport fra Havforskningen, 2021-7, 280 pp.
Gregory RS, Daborn GR (1982). Notes on adult lumpfish Cyclopterus lumpus L. from the Bay of Fundy. Proceedings of the Nova Scotian Institute of Science 32(4), 321–326.
Grotmol S, Totland GK, Thorud K, Hjeltnes BK (1997). Vacuolating encephalopathy and retinopathy associated with a nodavirus-like agent: a probable cause of mass mortality of cultured larval and juvenile Atlantic halibut Hippoglossus. Diseases of Aquatic Organisms 29: 85–97.
Guðmundsdóttir S, Vendramin N, Cuenca A, Sigurðardóttir H, Kristmundsson A, Moesgaard Iburg T, Jørgen Olesen N (2019). Outbreak of viral haemorrhagic septicaemia (VHS) in lumpfish (Cyclopterus lumpus) in Iceland caused by VHS virus genotype IV. Journal of Fish Diseases 42: 47–62. https://doi.org/10.1111/jfd.12910
Gwinn DC, Allen M, Johnston FD, Brown P, Todd CR, Arlinghaus R (2015). Rethinking length-based fisheries regulations: The value of protecting old and large fish with harvest slots. Fish and Fisheries 16: 259–281. https://doi.org/10.1111/faf.12053
Halvorsen KT (2017). Selective harvesting and life history variability of corkwing and goldsinny wrasse in Norway: Implications for management and conservation. Doctoral thesis. University of Oslo, Norway.
Halvorsen, KT, Larsen, T, Browman, HI, Durif, C, Aasen, N, Vøllestad, LA, Cresci A, Sørdalen TK, Bjelland R, Skiftesvik, AB. (2021). Movement patterns of temperate wrasses (Labridae) within a small marine protected area. Journal of Fish Biology, https://doi.org/10.1111/jfb.14825
Halvorsen KT, Larsen T, Sørdalen TK, Vøllestad LA, Knutsen H, Olsen EM (2017). Impact of harvesting cleaner fish for salmonid aquaculture assessed from replicated coastal marine protected areas. Marine Biology Research 13: 359–369. https://doi.org/10.1080/17451000.2016.1262042
Halvorsen KT, Sørdalen TK, Larsen T, Browman HI, Rafoss T, Albretsen J, Skiftesvik AB (2020a). Mind the depth: The vertical dimension of a small-scale fishery shapes selection on species, size and sex in wrasses. Marine and Coastal Fisheries, 12: 404-422. https://doi.org/10.1002/mcf2.10131
Halvorsen KT, Skiftesvik AB, Bjelland R, Larsen T (2020b). Kunnskapstøtte og råd for regulering av fisket etter leppefisk i 2021. English title: Management advice for Norwegian wrasse fisheries 2021. Rapport fra Havforskningen 51. Available from: https://www.hi.no/hi/nettrapporter/rapport-fra-havforskningen-2020-51
Halvorsen KT, Sørdalen TK, Durif C, Knutsen H, Olsen EM, Skiftesvik AB, Rustand TE, Bjelland RM, Vøllestad LA. (2016a). Male-biased sexual size dimorphism in the nest building corkwing wrasse (Symphodus melops): implications for a size regulated fishery. ICES Journal of Marine Science: Journal du Conseil 73: 2586–2594. https://doi.org/10.1093/icesjms/fsw135
Halvorsen KT, Sørdalen TK, Vøllestad LA, Skiftesvik AB, Espeland SH, Olsen EM (2016b). Sex- and size-selective harvesting of corkwing wrasse (Symphodus melops) - a cleaner fish used in salmonid aquaculture. ICES Journal of Marine Science: Journal du Conseil. 74(3):660–669. https://doi.org/10.1093/icesjms/fsw221
Hamilton SL, Caselle JE, Standish JD, Schroeder DM, Love MS, Rosales-Casian JA, Sosa-Nishizaki O (2007). Size-selective harvesting alters life histories of a temperate sex-changing fish. Ecological applications 17: 2268–2280. https://doi.org/10.1890/06-1930.1
Hansen KS, Steenberg C, Møller PR (2012). Small scale distribution of fish in offshore wind farms. How does renewable energy production affect aquatic life. ICES CM 2012/O: 11
Harkestad LS (2011). Eksperimentell smitte av grønngylt, Symphodus melops, med V. tapetis-isolatene CECT 4600, LP2 og NRP45. Mastergradsoppgave i havbruksbiologi, Universitetet i Bergen 2011, 136 s.
Hedeholm, R., M. E. Blicher and P. Gronkjaer (2014). First estimates of age and production of lumpsucker (Cyclopterus lumpus) in Greenland. Fisheries Research 149: 1–4. https://doi.org/10.1016/j.fishres.2013.08.016
Helland S, Dahle SW, Hough C, Borthen J (2014). Production of Ballan wrasse (Labrus bergylta). Science and Practice. The Norwegian Seafood Research Fund (FHF), p. 136.
Henly L, Stewart JE, Simpson SD (2021). Drivers and implications of change in an inshore multi-species fishery. ICES Journal of Marine Science, 78: 1815–1825. https://doi.org/10.1093/icesjms/fsab083
Herrmann B, Sistiaga M, Jørgensen T (2021). Size-dependent escape risk of lumpfish (Cyclopterus lumpus) from salmonid farm nets. Marine Pollution Bulletin 162: 111904.
Hilldén N-O (1984) Behavioural ecology of the labrid fishes (Teleostei, Labridae) at Tjärnö on the Swedish west coast. Doctoral dissertation, University of Stockholm, Sweden.
Hixon MA, Johnson DW, Sogard SM (2014). BOFFFFs: On the importance of conserving old-growth age structure in fishery populations. ICES Journal of Marine Science 71: 2171–2185. https://doi.org/10.1093/icesjms/fst200
Hjeltnes B, Bornø, G, Jansen, M D, Haukaas, A, Walde, C (Eds.) (2017) The Health Situation in Norwegian Aquaculture 2016 Norwegian Veterinary Institute https://www.vetinst.no/rapporter-og-publikasjoner/rapporter/2017/fish-health-report-2016/
Holm-Hansen TH, Carl H, Nielsen PG, Krag MA, Møller PR (2019). Assessing structure and seasonal variations of a temperate shallow water fish assemblage through Snorkel Visual Census. Cybium 43(4): 341–350. https://doi.org/10.26028/cybium/2019-434-004
Holst JC (1993). Observations on the distribution of lumpsucker (Cyclopterus lumpus, L.) in the Norwegian Sea. Fisheries Research, 17: 369–372.
Hvas M, Folkedal O, Imsland A, Oppedal F (2018). Metabolic rates, swimming capabilities, thermal niche and stress response of the lumpfish, Cyclopterus lumpus. Biology open, 7(9), bio036079. https://doi.org/10.1242/bio.036079
Hvas M, Folkedal O, Oppedal F (2020). Fish welfare in offshore salmon aquaculture. Reviews in Aquaculture 13, https://doi.org/10.1111/raq.12501
Imsland AK, Reynolds P, Eliassen G, Hangstad TA, Foss A, Vikingstad E, Elvegård TA (2014a). The use of lumpfish (Cyclopterus lumpus L.) to control sea lice (Lepeophtheirus salmonis Krøyer) infestations in intensively farmed Atlantic salmon (Salmo salar L.). Aquaculture 425-426: 18–23. https://doi.org/10.1016/j.aquaculture.2013.12.033
Imsland AK, Reynolds P, Eliassen G, Hangstad TA, Nytrø AV, Foss A, Vikingstad E, Elvegård TA (2014b). Notes on behaviour of lumpfish in sea pens with and without Atlantic salmon. Journal of Ethology 32: 117–122. https://doi.org/10.1007/s10164-014-0397-1
Imsland AK, Reynolds P, Eliassen G, Hangstad TA, Nytrø AV, Foss A, Vikingstad E, Elvegård TA (2014c). Assessment of growth and sea lice infection levels in Atlantic salmon stocked in small-scale cages with lumpfish. Aquaculture 433: 137–142. https://doi.org/10.1016/j.aquaculture.2014.06.008
Imsland AK, Reynolds P, Eliassen G, Hangstad TA, Nytrø AV, Foss A, Vikingstad E, Elvegård TA (2015a). Feeding preferences of lumpfish (Cyclopterus lumpus L.) maintained in open net-pens with Atlantic salmon (Salmo salar L.). Aquaculture 436: 47–51. https://doi.org/10.1016/j.aquaculture.2014.10.048
Imsland AK, Reynolds P, Eliassen G, Hangstad TA, Nytrø AV, Foss A, Vikingstad E, Elvegård TA (2015b). Assessment of suitable substrates for lumpfish in sea pens. Aquaculture International 23: 639–645. https://doi.org/10.1007/s10499-014-9840-0
Imsland AK, Reynolds P, Nytrø AA, Eliassen G, Hangstad TA, Jónsdóttir ÓDB, Emaus P-A, Elvegård TA, Lemmens SCA, Rydland R, Jonassen TM (2016a). Effects of lumpfish size on foraging behaviour and co-existence with sea lice infected Atlantic salmon in sea cages. Aquaculture 465: 19–27. https://doi.org/10.1016/j.aquaculture.2016.08.015
Imsland AK, Reynolds P, Eliassen G, Hangstad TA, Jónsdóttir ÓDB, Elvegård TA, Lemmens SCA, Rydland R, Nytrø AV (2016b). Investigation and quantification of behavioural interactions between lumpfish (Cyclopterus lumpus) and Goldsinny (Ctenolabrus rupestris) under controlled conditions. Aquaculture International 24: 1509–1521. https://doi.org/10.1007/s10499-016-0008-y
Imsland AK, Reynolds P, Eliassen G, Mortensen A, Hansen ØJ, Puvanendran V, Hangstad TA, Jónsdóttir ÓDB, Emaus PA, Elvegård TA, Lemmens SCA, Rydland R, Nytrø AV, Jonassen TM (2016c). Is cleaning behaviour in lumpfish (Cycloptherus lumpus) parentally controlled? Aquaculture 459: 156–165. https://doi.org/10.1016/j.aquaculture.2016.03.047
Imsland AKD, Hanssen A, Nytrø AV, Reynolds P, Jonassen TM, Hangstad TA, Elvegård TA, Urskog TC, Mikalsen B (2018). It works! Lumpfish can significantly lower sea lice infestation in large-scale salmon farming. Biology open 7(9): bio036301. doi.org/10.1242/bio.036301
Imsland AK, Reynolds P, Eliassen G, Berg-Hansen L, Hangstad, TA, Elvegård TA, Urskog TC, Mikalsen B. (2018b). Assessment of artificial substrates for lumpfish: effect of material thickness and water current speed. Aquaculture International 26, 1469-1479.
Imsland AKD, Reynolds P, Jonassen TM, Hangstad TA, Noble T, Wilson W, Mackie JA, Elvegård TA, Urskog TC, Mikalsen B (2019a). Comparison of feeding behaviour, growth and health of lumpfish (Cyclopterus lumpus L.) fed either feed blocks or pellets commercial feed. Aquaculture Research 50: 1952–1963. https://doi.org/10.1111/are.14083
Imsland AKD, Reynolds P, Jonassen TM, Hangstad TA, Elvegård TA, Urskog TC, Hanssen A, Mikalsen B (2019b). Effects of different feeding frequencies on growth, cataract development and histopathology of lumpfish (Cyclopterus lumpus L.). Aquaculture 501: 161–168. https://doi.org/10.1016/j.aquaculture.2018.11.026
Imsland AKD, Reynolds P, Lorentzen M, Eilertsen RA, Micallef G, Tvenning R. (2020). Improving survival and health of lumpfish (Cyclopterus lumpus L.) by the use of feed blocks and operational welfare indicators (OWIs) in commercial Atlantic salmon cages. Aquaculture 527, 735476. https://doi.org/10.1016/j.aquaculture.2020.735476
Ingólfsson A, Kristjánsson BK (2002). Diet of juvenile lumpsucker (Cyclopterus lumpus) in floating seaweed: Effect of ontogeny and prey availability. Copeia 2: 472–476.
Jansson E, Besnier F, Malde K, André C, Dahle G, Glover KA (2020). Genome wide analysis reveals genetic divergence between Goldsinny wrasse populations. BMC Genetics 21(1): 118. https://doi.org/10.1186/s12863-020-00921-8
Jansson E, Quintela M, Dahle G, Albretsen J, Knutsen H, André C, Strand Å, Mortensen S, Taggart JB, Karlsbakk E, Kvamme BO, Glover KA (2017). Genetic analysis of goldsinny wrasse reveals evolutionary insights into population connectivity and potential evidence of inadvertent translocation via aquaculture. ICES Journal of Marine Science 74: 2135–2147. https://doi.org/10.1093/icesjms/fsx046
Jensen Ø, Dempster T, Thorstad E, Uglem I, Fredheim A (2010). Escapes of fishes from Norwegian sea-cage aquaculture: causes, consequences and prevention. Aquaculture Environment Interactions, 1: 71–83. https://doi.org/10.3354/aei00008
Johansen R (ed.) (2013). Fiskehelserapporten 2012. Oslo, Veterinærinstituttet.
Johansen R, Tønnessen Ø, Nylund S (2019) Hvordan få kontroll på helseutfordringer på rensefisk. Foredrag Rensefiskkonferansen 2019.
Jonassen TM, Hamadi M, Remø SC, Waagbø R (2017). An epidemiological study of cataracts in wild and farmed lumpfish (Cyclopterus lumpus L.) and the relation to nutrition. Journal of Fish Diseases 40: 1903–1914. doi.org/10.1111/jfd.12664
Jonassen TM, Lein I, Nytrø AV (2018). Hatchery management of lumpfish. In: Treasurer JW (ed.). Cleaner fish biology and aquaculture applications. 5M Publications. Sheffield. pp. 122–146.
Jones M, Sommerville C, Wootten R (1992) Reduced sensitivity of the salmon louse, Lepeophtheirus salmonis, to the organophosphate dichlorvos. Journal of Fish Diseases 15:197-202. https://doi.org/10.1111/j.1365-2761.1992.tb00654.x
Jones S, Kim E, Bennett W (2008). Early development of resistance to the salmon louse, Lepeophtheirus salmonis (Krøyer), in juvenile pink salmon, Oncorhynchus gorbuscha (Walbaum). Journal of Fish Diseases 31: 591–600. https://doi.org/10.1111/j.1365-2761.2008.00933.x.
Jónsdóttir ÓDB, Schregel J, Hagen SB, Tobiassen C, Aarnes SG, Imsland AKD (2017). Population genetic structure of lumpfish along the Norwegian coast: aquaculture implications. Aquaculture International, 26(1): 49–60. doi.org/10.1007/s10499-017-0194-2
Karlsbakk E (2015). Amøbisk gjellesykdom (AGD) – litt om den nye plagen. Havforskningsrapporten 2015. Fisken og havet, særnr. 1-2015: 33–35.
Karlsbakk E, Olsen AB, Einen A-CB, Mo TA, Fiksdal IU, Aase H, Kalgraff C, Skår S-Å, Hansen H (2013). Amoebic gill disease due to Paramoeba perurans in Ballan wrasse (Labrus bergylta). Aquaculture 412-413, 41–44. https://doi.org/10.1016/j.aquaculture.2013.07.007
Kennedy J, Jónsson SÞ, Kasper JM, Olafsson HG 2015. Movements of female lumpfish (Cyclopterus lumpus) around Iceland. ICES Journal of Marine Science 72:880–889. doi.org/10.1093/icesjms/fsu170
Kennedy J, Jónsson SÞ, Ólafsson HG, Kasper JM 2016. Observations of vertical movements and depth distribution of migrating female lumpfish (Cyclopterus lumpus) in Iceland from data storage tags and trawl surveys. ICES Journal of Marine Science 73:1160–1169. doi.org/10.1093/icesjms/fsv244
Kennedy J (2018). Oocyte size distribution reveals ovary development strategy, number and relative size of egg batches in lumpfish (Cyclopterus lumpus). Polar Biology 41(6): 1091–1103. https://doi.org/10.1007/s00300-018-2266-9
Kennedy J, Durif CMF, Florin A-B, Fréche A, Gauthier J, Hüssy K, Jónsson SÞ, Ólafsson HG, Post S, Hedeholm RB (2019). A brief history of lumpfishing, assessment, and management across the North Atlantic. ICES Journal of Marine Science 76(1): 181–191. https://doi.org/10.1093/icesjms/fsy146
Kindsvater HK, Mangel M, Reynolds JD, Dulvy NK (2016). Ten principles from evolutionary ecology essential for effective marine conservation. Ecology and Evolution 6: 2125–2138. https://doi.org/10.1002/ece3.2012
Kindsvater HK, Halvorsen, KT, Sørdalen, TK, Alonzo SH (2020). The consequences of size‐selective fishing mortality for larval production and sustainable yield in species with obligate male care. Fish and Fisheries 21: 1135–1149. https://doi.org/10.1111/faf.12491
Klakegg O, Myhren S, Juell RA, Aase M, Salonius K, Sørum H (2020). Improved health and better survival of farmed lumpfish (Cyclopterus lumpus) after a probiotic bath with two probiotic strains of Aliivibrio. Aquaculture 518: 734810. https://doi.org/10.1016/j.aquaculture.2019.734810
Kleiven, AR, SH Espeland, G Søvik, J Albretsen, PJ Nillos Kleiven, F Zimmerman, Grefsrud ES, KT Halvorsen, O Vie (2021). Aktiv forvaltning av marine ressurser - Frøya og Hitra. Rapport fra Havforskningen (14). https://www.hi.no/en/hi/nettrapporter/rapport-fra-havforskningen-2021-14
Knausgård, K. M., Wiklund, A., Sørdalen, T. K., Halvorsen, K., Kleiven, A. R., Jiao, L., & Goodwin, M. (2020). Temperate FishDetection and Classification: a Deep Learning based Approach. Applied Intelligence. https://doi.org/10.1007/s10489-020-02154-9
Knutsen H, Jorde PE, Gonzalez EB, Robalo J, Albretsen J, Almada V, (2013). Climate Change and Genetic Structure of Leading Edge and Rear End Populations in a Northwards Shifting Marine Fish Species, the Corkwing Wrasse (Symphodus melops). PLoS ONE 8, e67492. https://doi.org/10.1371/journal.pone.0067492
Korsnes K, Karlsbakk E, Skaar CK, Sælemyr L, Nylund A, Kvamme BO, Mortensen S (2017). High nervous necrosis virus (NNV) diversity in wild wrasse (Labridae) in Norway. Diseases of Aquatic Organisms 126, 43–50. https://doi.org/10.3354/dao03159
Kousoulaki K, Migaud H, Davie A (2018). Cleaner fish species nutrition and feeding practices. In: Treasurer JW (ed.). Cleaner fish biology and aquaculture applications. 5M Publications. Sheffield. pp. 179–196.
Kristiansen TS, Fernö A, Pavlidis MA, van de Vis H (2020). The Welfare of Fish. Springer Nature Switzerland. 515 pp. https://doi.org/10.1007/978-3-030-41675-1
Kvenseth PG (1996). “Large-scale use of wrasse to control sea lice and net fouling in salmon farms in Norway,” in Proceedings of the Wrasse Biology and Aquaculture Applications, eds M. D. J. Sayer, J. W. Treasurer, and M. J. Costello (Oban: Blackwell), 196–203.
Laikre L, Schwartz MK, Waples RS, Ryman N, GeM Working Group (2010). Compromising genetic diversity in the wild: Unmonitored large-scale release of plants and animals. Trends in Ecology and Evolution, 25(9), 520–529. https://doi.org/10.1016/j.tree.2010.06.013
Le H.T. M D., Shao, Xi., Krogdahl, Å., Kortner, T.M., Lein, I., Kousoulaki, K., Lie, K. K., (2019). Intestinal Function of the Stomachless Fish, Ballan Wrasse (Labrus bergylta). Frontiers in Marine Science 6: 1-15. https://doi.org/10.3389/fmars.2019.00140
Lees F, Baillie M, Gettinby G, Revie CW (2008). The efficacy of emamectin benzoate against infestations of Lepeophtheirus salmonis on farmed Atlantic salmon (Salmo salar L) in Scotland, 2002–2006. PLoS ONE 3(2), e1549. doi:10.1371/journal.pone.0001549.
Lie KK, Tørresen OK, Solbakken MH, Rønnestad I, Tooming-Klunderud A, Nederbragt AJ, Jentoft S, Sæle Ø (2018). ‘Loss of Stomach, Loss of Appetite? Sequencing of the Ballan Wrasse (Labrus bergylta) Genome and Intestinal Transcriptomic Profiling Illuminate the Evolution of Loss of Stomach Function in Fish’. BMC Genomics 19, no. 1 (6 March 2018): 186. https://doi.org/10.1186/s12864-018-4570-8
Loew E (2012). Kartlegging av hvilke lyskvaliteter berggylt er i stand til å se i fiskens ulike livsstadier. Rapport fra Havforskningen, Nr 28, 2012.
Lusedata.no Salmon louse statistics. https://lusedata.no/
Matić-Skoko S, Bojanić Varezić D., Šiljić J, Tutman P, Pallaoro A (2013). The cuckoo wrasse, Labrus mixtus (Pisces: Labridae): biological indices for life history and conservation. Scientia Marina, 77(4), 595–605. http://doi.org/10.3989/scimar.03884.07A
Maunder MN, Punt AE (2004). Standardizing catch and effort data: a review of recent approaches. Fisheries Research 70, 141–159.
MarinHelse (2018). Veileder i Dødfiskkategorisering. 101pp. Developed by MarinHelse AS, Tromsø, Norway. https://www.marinhelse.no/produkter/veileder-dodfiskkategorisering/
Maroni K, Andersen P (1996). Distribution and abundance of wrasse in an area of northern Norway. In: Sayer MDJ, Costello MJ, Treasurer JW (Eds.). Wrasse: Biology and use in Aquaculture. (pp. 70–73). Fishing News Books.
Mattingsdal, M., Jentoft, S., Tørresen, O. K., Knutsen, H., Hansen, M. M., Robalo, J. I., … Gonzalez, E. B. (2018). A continuous genome assembly of the corkwing wrasse (Symphodus melops). Genomics, 110, 399–403.
Mattingsdal M, Jorde PE, Knutsen H, Jentoft S, Stenseth NC, Sodeland M, Robalo JI, Hansen MM, André C, Gonzalez EB (2020). Demographic history has shaped the strongly differentiated corkwing wrasse populations in Northern Europe. Molecular Ecology 29: 160–171. https://doi.org/10.1111/mec.15310
Ministry of Trade, Industry and Fisheries (2017). Regulation on production areas for grow-out of salmon, trout and Rainbow trout in Sea (in Norwegian: Forskrift Om Produksjonsområder for Akvakultur Av Matfisk i sjø av laks, ørret og regnbueørret (Produksjonsområdeforskriften)), FOR-2017-01-16-6, lovdata.no, https://lovdata.no/dokument/SF/forskrift/2017-01-16-61
Ministry of Trade, Industry and Fisheries (2012). Regulation on combatting salmon lice in aquaculture facilities (in Norwegian: Forskrift om bekjempelse av lakselus i akvakulturanlegg), FOR-2012-12-05-1140, lovdata.no, https://lovdata.no/dokument/SF/forskrift/2012-12-05-1140
Ministry of Trade, Industry and Fisheries (2021). A sea of possibilities – the government's aquaculture strategy (in Norwegian: Et hav av muligheter – regjeringens havbruksstrategi. Ministry of Trade, Industry and Fisheries, Norway.
Mitamura H, Thorstad EB, Uglem I, Bjorn PA, Okland F, Naesje TF, Dempster T, Arai N (2012). Movements of lumpsucker females in a northern Norwegian fjord during the spawning season. Environmental Biology of Fishes 93(4): 475–481. https://doi.org/10.1007/s10641-011-9942-8
Mo TA, Emilsen V, Hansen H, Poppe, TT (2014). Smitteforsøk med isolat av Paramoeba perurans fra berggylt og laks for å studere mottakelighet og infeksjonsutvikling hos laks. Rapport, Veterinærinstituttet 31 Jan. 2014. 13 s.
Moland, E., Fernández-Chacón, A., Sørdalen, T. K., Villegas-Ríos, D., Thorbjørnsen, S. H., Halvorsen, K. T., … Knutsen, J. A. (2021). Restoration of Abundance and Dynamics of Coastal Fish and Lobster Within Northern Marine Protected Areas Across Two Decades. Frontiers in Marine Science, 8, 1–12.
Mortensen S, Skiftesvik AB, Bjelland R, Karlsbakk E, Durif C, Sandlund N (2017). Bruk av rensefisk i lakseoppdrett. pp 162-173. In: Svåsand T, Grefsrud ES, Karlsen Ø, Kvamme BO, Glover KS, Husa V, Kristiansen TS (eds.). Risikorapport norsk fiskeoppdrett 2017. Fisken og havet, særnummer 2 – 2017. 179 pp (in Norwegian).
Muncaster S, Andersson E, Kjesbu OS, Taranger GL, Skiftesvik AB, Norberg B (2010). The reproductive cycle of female Ballan wrasse Labrus bergylta in high latitude, temperate waters. Journal of Fish Biology 77: 494-511. https://doi.org/10.1111/j.1095-8649.2010.02691.x
Muncaster S, Norberg B, Andersson, E (2013). Natural sex change in the temperate protogynous Ballan wrasse Labrus bergylta. Journal of Fish Biology 82: 1858–1870. https://onlinelibrary.wiley.com/doitemanord2021-545.pdf10.1111/jfb.12113
Munro ES, McIntosh RE, Weir SJ, Noguera PA, Sandilands JM, Matejusova I, Mayes AS, Smith, R (2015). A mortality event in wrasse species (Labridae) associated with the presence of viral haemorrhagic septicaemia virus. Journal of Fish Diseases 38: 335–341. doi.org/10.1111/jfd.12237
Munro L, Wallace I (2018) Marine Scotland Science—Scottish shellfish farm production survey 2017. Aberdeen: Marine Scotland Science
Nedreaas K, Aglen A, Gjøsæter J, Jørstad K, Knutsen H, Smedstad O, Svåsand T, Agotnes P (2008). Management of cod in Western Norway and on the Skagerrak coast – stock status and possible management measures. Fisken og Havet, 5: 1–106.
Nilsen A, Viljugrein H, Rosag MV, Colquhon D (2014). Rensefiskhelse - kartlegging av dodelighetssarsaker. Veterinærinstituttets rapportserie 12. In Norwegian.
Noble C, Iversen MH, Lein I, Kolarevic J, Johansen L–H, Burgerhout E, Puvanendran V, Kousoulaki K, Aas GH, Stene A, Espmark ÅM (2019a). RENSVEL OWI FACT SHEET SERIES: An introduction to Operational and Laboratory based Welfare Indicators for Ballan wrasse (Labrus bergylta). 43 pp. ISBN 978-82-8296-590-3
Noble C, Iversen MH, Lein I, Kolarevic J, Johansen L-H, Berge GM, Burgerhout E, Puvanendran V, Mortensen A, Stene A, Espmark ÅMO (2019). RENSVEL OWI FACT SHEET SERIES: An introduction to Operational and Laboratory-based Welfare Indicators for lumpfish (Cyclopterus lumpus L.). Research report. 46 pp. Available from: https://ntnuopen.ntnu.no/ntnu-xmlui/handle/11250/2599803
NFSA (2020). Sluttrapport: Nasjonalt tilsynskampanje 2018/2019: Velferd hos rensefisk. The Norwegian Food Authorities, Norway https://www.mattilsynet.no/fisk_og_akvakultur/akvakultur/rensefisk/rensefiskkampanjen_20182019.30882
Nytrø AV, Vikingstad E, Foss A, Hangstad TA, Reynolds P, Eliassen G, Elvegård TA, Falk-Pedersen I-B, Imsland AK (2014). The effect of temperature and fish size on growth of juvenile lumpfish (Cyclopterus lumpus L.). Aquaculture 434: 296–302. https://doi.org/10.1016/j.aquaculture.2014.07.028
Olsen EM, Halvorsen KT, Larsen T, Kuparinen A (2018). Potential for managing life history diversity in a commercially exploited intermediate predator, the goldsinny wrasse (Ctenolabrus rupestris). ICES Journal of Marine Science 76(2): 410–417. https://fishlarvae.org/wp-content/uploads/2018/07/Olsen-et-al..pdf
Overton K, Barrett LT, Oppedal F, Kristiansen TS, Dempster T (2020). Sea lice removal by cleaner fish in salmon aquaculture: a review of the evidence base; Aquaculture Environmental Interactions 12: 31–44. https://doi.org/10.3354/aei00345
Pampoulie C, Skirnisdottir S, Olafsdottir G, Helyar SJ, Thorsteinsson V, Jónsson SÞ, Fréchet A, Durif CMF, Sherman S, Lampart-Kałużniacka M, Hedeholm R, Ólafsson H, Daníelsdóttir, AK, Kasper JM (2014). Genetic structure of the lumpfish Cyclopterus lumpus across the North Atlantic. ICES Journal of Marine Science, 71(9): 2390–2397. https://doi.org/10.1093/icesjms/fsu071
Patel S, Korsnes K, Bergh Ø, Vik-Mo F, Pedersen J, Nerland AH (2007). Nodavirus in farmed Atlantic cod Gadus morhua in Norway. Diseases of Aquatic Organisms 77: 169–173. https://doi.org/10.3354/dao01842
Peeler EJ, Feist SW (2011). Human intervention in freshwater ecosystems drives disease emergence. Freshwater Biology 56(4): 705–716. doi.org/10.1111/j.1365-2427.2011.02572.x
Peeler EJ, Oidtmann BC, Midtlyng PJ, Miossec L, Gozlan RE (2011). Non-native aquatic animals introductions have driven disease emergence in Europe. Biological Invasions 13(6): 1291–303. doi.org/10.1007/s10530-010-9890-9
Poppe TT, Taksdal T, Skjelstad H, Sviland C, Vågnes Ø, Colquhoun DJ (2012). Nye arter – nye diagnostiske utfordringer. Norsk Veterinærtidsskrift 124(1): 19–21.
Potts GW (1974). The colouration and its behavioural significance in the corkwing wrasse, Crenilabrus Melops. Journal of the Marine Biological Association of the United Kingdom 94: 925–938.
Potts GW (1985). The nest structure of the corkwing wrasse Crenilabrus melops (Labridae, Teleostei). Journal of the Marine Biological Association of the United Kingdom, 65: 531 –546.
Powell A, Pooley C, Scolamacchia M, Garcia de Leaniz C (2018a). Review of lumpfish biology. In J. W. Treasurer (Ed.), Cleaner fish biology and aquaculture applications (pp. 98–121). 5M Publishing Ltd.
Powell, A., J. W. Treasurer, C. L. Pooley, A. J. Keay, R. Lloyd, A. K. Imsland and C. G. de Leaniz (2018b). Use of lumpfish for sea-lice control in salmon farming: challenges and opportunities. Reviews in Aquaculture 10(3): 683-702. https://doi.org/10.1111/raq.12194
Putman AI, Carbone I (2014). Challenges in analysis and interpretation of microsatellite data for population genetic studies. Ecology and Evolution 4(22): 4399–4428. doi:10.1002/ece3.1305
Quintela M, Danielsen EA, Lopez L, Barreiro R, Svåsand T, Knutsen H, Skiftesvik, AB, Glover KA (2016). Is the ballan wrasse (Labrus bergylta) two species? Genetic analysis reveals within-species divergence associated with plain and spotted morphotype frequencies. Integrative Zoology 11: 162–172. https://doi.org/10.1111/1749-4877.12186
Rabadan CC, Spreadbury C, Consuegra C, Garcia de Leaniz C (2021). Development, validation and testing of an Operational Welfare Score Index forfarmed lumpfish Cyclopterus lumpus L. Aquaculture 531. 1-10. https://doi.org/10.1016/j.aquaculture.2020.735777
Rae GH (2002). Sea louse control in Scotland, past and present. Pest Management Science: formerly Pesticide Science 58:515-520. https://doi.org/10.1002/ps.491
Reamon, M (2020). Assessing the impacts of marine protected areas on wrasse populations in Norway. Master thesis. University of Agder.
Reiss H, Hoarau G, Dickey‐Collas M, Wolff WJ (2009). Genetic population structure of marine fish: mismatch between biological and fisheries management units. Fish and Fisheries, 10: 361–395. doi.org/10.1111/j.1467-2979.2008.00324.x
Rieber-Mohn GF, Poppe TT, Mo TA (2021). En illusorisk dyrevelferd, Norsk Veterinærtidsskrift nr 1, 2021.https://nvt.vetnett.no/journal/2021/1/m-51/En_illusorisk_dyrevelferd. In Norwegian
Robalo JI, Castilho R, Francisco SM, Almada F, Knutsen H, Jorde PE, Pereira AM, Almada V (2012). Northern refugia and recent expansion in the North Sea: the case of the wrasse Symphodus melops (Linnaeus, 1758). Ecology and Evolution 2: 153–164. https://doi.org/10.1002/ece3.77
Roth M, Richards RH, Sommerville C (1993). Current practices in the chemotherapeutic control of sea lice infestations in aquaculture: a review. Journal of Fish Diseases 16: 1-26.
Rui Beja P (1995). Structure and Seasonal Fluctuations of Rocky Littoral Fish Assemblages in South-Western Portugal: Implications for Otter Prey Availability. Journal of the Marine Biological Association of the United Kingdom 75: 833.
Rådet for dyreetikk (2020). Er det mulig å bruke rensefisk på en etisk og velferdsmessig forsvarlig måte i norske oppdrettsanlegg? https://www.radetfordyreetikk.no/er-det-mulig-a-bruke-rensefisk-pa-en-etisk-og-velferdsmessig-forsvarlig-mate-i-norske-oppdrettsanlegg/?fbclid=IwAR3UT83VqXGRvYEr5nF9PyHkcAJ9_rqwiTRDhY-nmMHuPci_4O9FcDPNb44 (In Norwegian)
Samuelsen T, Vik BL, Standal O (2020). Alle snakker om bærekraft, men ingen snakker om hva det betyr. Kronikk, Universiteter I Agder, Universietsavisa (https://www.universitetsavisa.no/alle-snakker-om-baerekraft-men-ingen-snakker-om-hva-det-betyr/113877)
Sandlund N, Mortensen S, Nedreaas K, Skiftesvik AB, Jansson E, Halvorsen K (2020). Kunnskapsstatus: Miljøeffekter ved bruk av villfanget leppefisk i fiskeoppdrett, Kap 6, https://www.hi.no/resources/kunnskapsstatus-risikorapportar/2019/Kap_6_Miljoeffekter-ved-bruk-av-villfanget-leppefisk-i-fiskeoppdrett.pdf In Norwegian.
Sandlund N, Rønneseth A, Ellul RM, Nylund S, Sandlund L (2021). Pasteurella spp. Infections in Atlantic salmon and lumpsucker. Journal of Fish Diseases.;n/a(n/a).
Sayer MDJ (1999). Duration of refuge residence by goldsinny, Ctenolabrus rupestris. Journal of the Marine Biological Association of the United Kingdom 79: 571–572.
Sayer MDJ, Davenport J (1996). Hypometabolism in torpid goldsinny wrasse subjected to rapid reductions in seawater temperature. Journal of Fish Biology 49: 64–75. doi.org/10.1111/j.1095-8649.1996.tb00005.x
Sayer MDJ, Gibson RN, Atkinson RJA (1995). Growth, diet and condition of goldsinny on the west coast of Scotland. Journal of Fish Biology 46: 317–340.
Sayer MDJ, Gibson RN, Atkinson RJA (1996a). The biology of inshore goldsinny populations: can they sustain commercial exploitation? Pp 91–99. In: Sayer MDJ, Costello MJ, Treasurer JW (eds). Wrasse Biology and use in Aquaculture. Oxford: Fishing News Books,
Sayer MDJ, Gibson RN, Atkinson RJA (1996b). Growth, diet and condition of corkwing wrasse and rock cook on the west coast of Scotland. Journal of Fish Biology 49: 76–94.
Sayer, M. D. J., Gibson, R. N., & Atkinson, R. J. A. (1993). Distribution and density of populations of goldsinny wrasse (Ctenolabrus rupestris) on the west coast of Scotland. Journal of Fish Biology, 43, 157–167.
Sayer MDJ, Reader JP (1996). Exposure of goldsinny, rock cook and corkwing wrasse to low temperature and low salinity: Survival, blood physiology and seasonal variation. Journal of Fish Biology 49: 41-63.
Schönherz AA, Hansen MHH, Jorgensen HBH, Berg P, Lorenzen N, Einer-Jensen K (2012). Oral transmission as a route of infection for viral haemorrhagic septicaemia virus in rainbow trout, Oncorhynchus mykiss (Walbaum). Journal of Fish Diseases 35: 395-406. https://doi.org/10.1111/j.1365-2761.2012.01358.x
Seljestad G, Quintela M, Faust E, Halvorsen KT, Besnier F, Jansson E, Dahle G, Knutsen H, André C, Folkvord A, Glover KA (2020). “A cleaner-break”: Genetic divergence between geographic groups and sympatric phenotypes revealed in Ballan wrasse (Labrus bergylta). Ecology and Evolution 10: 6120–6135. https://doi.org/10.1002/ece3.6404
Sevatdal S, Horsberg TE (2003) Determination of reduced sensitivity in sea lice (Lepeophtheirus salmonis Krøyer) against the pyrethroid deltamethrin using bioassays and probit modelling. Aquaculture 218: 21-31. https://doi.org/10.1016/S0044-8486(02)00339-3
Sevatdal S, Copley L, Wallace C, Jackson D, Horsberg TE (2005). Monitoring of the sensitivity of sea lice (Lepeophtheirus salmonis) to pyrethroids in Norway, Ireland and Scotland using bioassays and probit modelling. Aquaculture 244: 19–27.
Sigsgaard EE, Nielsen IB, Carl H, Krag MA, Knudsen SW, Xing Y, Holm‑Hansen TH, Møller PR, Thomsen PF (2017). Seawater environmental DNA reflects seasonality of a coastal fish community. Marine Biology 164:128. https://doi.org/10.1007/s00227-017-3147-4
Skiftesvik AB, Boxaspen K, Parsons A (1996). Preliminary breeding trials of wrasse in an intensive system. In Wrasse: Biology and Use in Aquaculture (Sayer MDJ, Treasurer JW, Costello MJ (eds), pp. 136–141. Oxford: Blackwell Scientific.
Skiftesvik AB, Bjelland RM, Durif CMF, Johansen IS, Browman HI (2013). De-lousing of Atlantic salmon (Salmo salar) by cultured vs. wild Ballan wrasse (Labrus bergylta). Aquaculture 402-403: 113-118. https://doi.org/10.1016/j.aquaculture.2013.03.032
Skiftesvik AB, Durif CMF, Bjelland RM, Browman HI (2014). Distribution and habitat preferences of five species of wrasse (family Labridae) in a Norwegian fjord. ICES Journal of Marine Science 72: 890–899. https://doi.org/10.1093/icesjms/fsu211
Skog K, Mikkelsen KO, Bjordal Å (1994). Leppefisk – Tilgjengelighet og fangstmuligheter i perioden februar - juni. Fisken og Havet nr 4, 1–34. In Norwegian.
Skoge RH, Brattespe J, Økland AL, Plarre H, Nylund A, (2017). New virus of the family Flaviviridae detected in lumpfish (Cyclopterus lumpus). Archives of Virology 163: 679–685. https://doi.org/10.1007/s00705-017-3643-3
Sommerset I, Walde C S, Bang Jensen B, Bornø B, Haukaas A og Brun E (red). (2020) Fiskehelserapporten 2019, Oslo, Veterinærinstituttet
Stagg HEB, Guðmundsdóttir S, Vendramin N, Ruane NM, Sigurðardóttir H, Christiansen DH, Cuenca A, Petersen PE, Munro ES, Pobov VL, Subramaniam K, Imnoi K, Waltzek TB,
Olesen NJ (2020). Characterization of ranaviruses isolated from lumpfish Cyclopterus lumpus L. in the North Atlantic area: proposal for a new ranavirus species (European North Atlantic Ranavirus). Journal of General Virology 101(2): 198-207. https://doi.org/10.1099/jgv.0.001377
Stien LH, Størkesen KV, Gåsnes SK (2020). «Analyse av dødelighetsdata fra spørreundersøkelse om velferd hos rensefisk». Rapport fra Havforskningen, 2020–6. The Institute of Marine Research, Norway.
Stone J (1996). Preliminary trials on the culture of goldsinny and corkwing wrasse. In Wrasse: Biology and Use in Aquaculture (Sayer, M. D. J., Treasurer, J. W. & Costello, M. J., eds), pp. 142–167. Oxford: Blackwell Scientific
Steven GA (1933). The food consumed by shags and cormorants around the shores of Cornwall (England). Journal of the Marine Biological Association of the United Kingdom 19: 277.
Størkersen KV, Amundsen VS (2019). Rensefiskens ve og vel i merdene. Resultat av spørreundersøkelse til matfisklokaliteter. NTNU Samfunnsforskning, Studio Apertura, Norway.
Støttrup JG, Kokkalis A, Brown EJ, Olsen J, Andersen SK, Pedersen EM (2018). Harvesting geo-spatial data on coastal fish assemblages through coordinated citizen science. Fisheries Research 208: 86-96. https://doi.org/10.1016/j.fishres.2018.07.015
St&
Appendix 1
Geographical distribution of Norwegian retained catches of wild wrasse species in 2019

Appendix figure 1. Geographical distribution of reported Norwegian retained catches of the different wrasse species in 2019 per statistical areas. The grey-shaded area shows the area inside 12 nautical miles from the baseline. Note the different colour scales. The horizontal blue lines mark the borders between the three management areas.
Appendix 2
Key points in the Norwegian wrasse fishery regulations for 2020
Position reporting
From 2020, a requirement will be introduced for participating vessels to be equipped with type-approved automatic identification system (AIS, Class A or B) or VMS. This requirement applies to all fishermen who engage in commercial fishing, incl. recreational fishers. The requirement for position reporting is introduced as a consequence of the need to be able to control the fishing activities effectively. In addition, position reports provide more precise knowledge about where the fishing is going on. This is something that provides a better basis for the management of wrasses and the setting of quota levels.
Submission of a delivery agreement
In order to participate in the fishery for wrasses, one of the conditions for participation is that there must be a delivery agreement with an approved buyer.
Anyone who is to participate in wrasse fishing must submit a delivery agreement to the Directorate of Fisheries before the vessel starts fishing for wrasses.
Permission to use fyke nets
Along the coast from the border with Sweden to the Møre and Romsdal county, it is forbidden to use fyke nets from 1 May to 31 December. Professional fishermen who may apply for a license to use fyke nets shall use the same form as when submitting a delivery agreement.
Total Quota
- The total quota of 18 million wrasses is divided into three regions:
- 4 million wrasses on the coast from the border with Sweden to Varnes lighthouse on Lista (southern area)
- 10 million wrasses on the coast from Varnes lighthouse on Lista to 62 degrees north (western area)
- 4 million wrasses on the coast north of 62 degrees north (northern area)
- The total quota is distributed by 90 per cent to a closed group of fishers and 10 per cent to an open group.
Closed group
- Vessel quotas in the closed group are 48 000 wrasses per vessel.
Open group
- Maximum open group quotas are 5000 wrasses per vessel.
- For participation in the open group, general terms and conditions apply. It is among other conditions that the owner himself is the master on board the vessel, unless an exemption has been granted in accordance with general rules.
- Furthermore, vessel owners can only participate with one vessel in the open group.
- Vessel owners who participate with vessels in the closed group cannot also participate with vessels in the open group.
Recreational fishing
- It will still be possible to apply for a license for recreational fishing for wrasses for sale under certain conditions.
If the Director of Fisheries stops fishing in an open group, there will also be a stop in the recreational fishing for wrasses.
Gear restrictions
- On the coastline from the Swedish border to the Varnes lighthouse on Lista (N 58˚10.7’): 100 traps or fyke nets per vessel owner
- On the coastline from Varnes lighthouse on Lista and northwards: 400 traps or fyke nets per vessel owner
There is a requirement to use entry barriers in the gear to reduce by-catch of species other than wrasses, although the fishers are required to immediately release all by-catch back into the sea.
From 2021 onwards, a requirement will be that the fish entrance in wrasse traps should have a diameter of 60 mm or less.
There is also a requirement to use escapement grids (with minimum 12 mm between the grid bars) to ensure that the smallest wrasses can easily escape from the gear.
Phasing out fyke nets
From 2020 it is forbidden for recreational fishers to use fyke nets to catch wrasses
For professional fishermen who fish with vessels registered in the national fishing vessel registry, it will be forbidden to use fyke nets to catch wrasses from 2021 onwards.
Bycatch
It is not allowed to keep by-catches in traps or fyke nets that are used to catch wrasses. This also applies to the by-catch of crabs. All by-catch should immediately be returned to the sea in such a way that it is not damaged and that it returns to its natural environment. This rule also applies to wrasse species caught in violation of the current minimum size regulation or which for other reasons must be released back into the sea.
Fishing periods
- South of 62˚N: from July 17 at 08:00 to October 20 at 20:00
- North of 62˚N: from July 31 at 08:00 to October 20 at 20:00
About this publication
Towards a sustainable fishery and use of cleaner fish in salmonid aquaculture
Challenges and opportunities
Kim Tallaksen Halvorsen, Anne Berit Skiftesvik, Caroline Durif, Ellika Faust, Håkan Wennhage, Carl André, Jacob Linnemann Rønfeldt, Peter Rask Møller, Henrik Carl, Terje Jørgensen, Maria Quintela, Nina Sandlund, Lars Helge Stien, Kjell Nedreaas, Eeva Jansson, Hans Hagen Stockhausen, Kjetil Korsnes, Patrick Reynolds, Albert Imsland, Inger Fyllingen, Hulda Bysheim, Lauren Henly, Stein Mortensen
ISBN 978-92-893-7181-0 (PDF)
ISBN 978-92-893-7182-7 (ONLINE)
http://dx.doi.org/10.6027/temanord2021-545
TemaNord 2021:545
ISSN 0908-6692
© Nordic Council of Ministers 2021
Cover photo: Erling Svendsen
All illustrations are made by Stein Mortensen
Published: 3/12/2021
Disclaimer
This publication was funded by the Nordic Council of Ministers. However, the content does not necessarily reflect the Nordic Council of Ministers’ views, opinions, attitudes or recommendations.
Rights and permissions
This work is made available under the Creative Commons Attribution 4.0 International license (CC BY 4.0) https://creativecommons.org/licenses/by/4.0.
Translations: If you translate this work, please include the following disclaimer: This translation was not produced by the Nordic Council of Ministers and should not be construed as official. The Nordic Council of Ministers cannot be held responsible for the translation or any errors in it.
Adaptations: If you adapt this work, please include the following disclaimer along with the attribution: This is an adaptation of an original work by the Nordic Council of Ministers. Responsibility for the views and opinions expressed in the adaptation rests solely with its author(s). The views and opinions in this adaptation have not been approved by the Nordic Council of Ministers.
Third-party content: The Nordic Council of Ministers does not necessarily own every single part of this work. The Nordic Council of Ministers cannot, therefore, guarantee that the reuse of third-party content does not infringe the copyright of the third party. If you wish to reuse any third-party content, you bear the risks associated with any such rights violations. You are responsible for determining whether there is a need to obtain permission for the use of third-party content, and if so, for obtaining the relevant permission from the copyright holder. Examples of third-party content may include, but are not limited to, tables, figures or images.
Photo rights (further permission required for reuse):
Any queries regarding rights and licences should be addressed to:
Nordic Council of Ministers/Publication Unit
Ved Stranden 18
DK-1061 Copenhagen
Denmark
pub@norden.org
Nordic co-operation
Nordic co-operation is one of the world’s most extensive forms of regional collaboration, involving Denmark, Finland, Iceland, Norway, Sweden, and the Faroe Islands, Greenland and Åland.
Nordic co-operation has firm traditions in politics, economics and culture and plays an important role in European and international forums. The Nordic community strives for a strong Nordic Region in a strong Europe.
Nordic co-operation promotes regional interests and values in a global world. The values shared by the Nordic countries help make the region one of the most innovative and competitive in the world.
The Nordic Council of Ministers
Nordens Hus
Ved Stranden 18
DK-1061 Copenhagen
pub@norden.org
Read more Nordic publications on www.norden.org/publications